前言 B 细胞具有产生针对多种靶标的抗体的独特能力,可提供针对感染的保护,同时还有助于免疫失调环境中的发病机制。人类 B 细胞分为五个群体:过渡、幼稚、非转换记忆、转换记忆和浆细胞。识别和分类人类 B 细胞的功能亚群,阻碍了作者在自身免疫中选择性靶向致病性 B 细胞和在疫苗接种中诱导记忆反应的能力。
为了表征外围成熟的人类 B 细胞,本文作者开发了一种高度复用的单细胞筛选方案,通过使用大规模细胞术来量化 351 个表面分子的共表达。基于作者的研究结果,作者提出了一种分类方案,将来自外周血、骨髓、淋巴结和扁桃体四个组织的的 B 细胞分为 12 个独特的亚组,并构建了具有表面表型、代谢、生物合成活性和对免疫激活的信号反应特征的广泛单细胞图谱。这个人类 B 细胞身份图谱将使研究能够在稳态、疫苗接种、感染、自身免疫和癌症的背景下进一步确定 B 细胞亚群的功能。
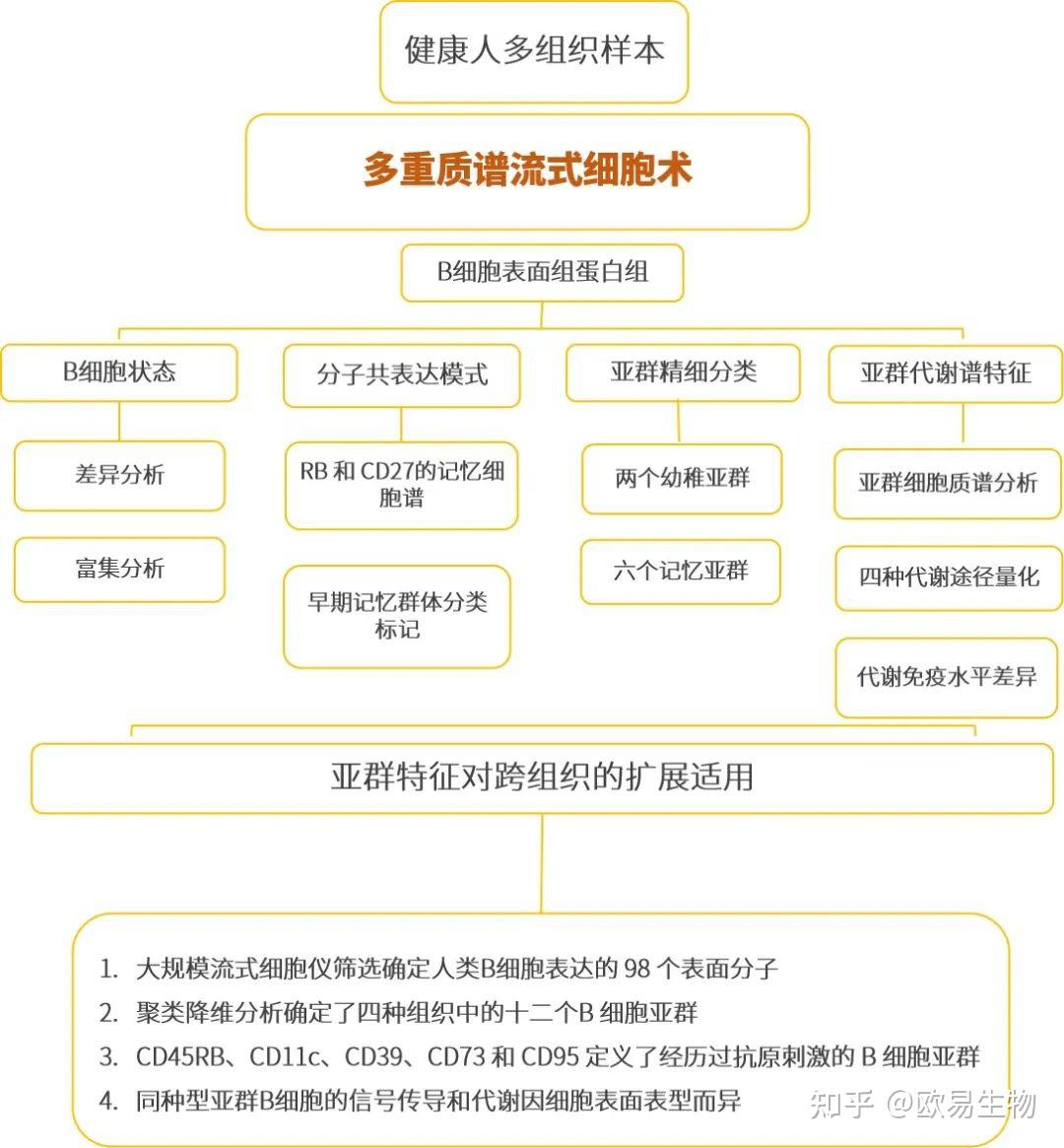
本篇为斯坦福大学研究团队在 Immunity期刊(IF:43.474)发表的题为 “An Integrated Multi-omic Single-Cell Atlas of Human B Cell Identity”的研究成果,采用改进的质谱流式细胞仪、流式细胞术等研究方法,成功量化了百万级人类B 细胞上 351 种表面分子的共表达模式。通过鉴定了差异表达的分子,对比VDJ 序列、代谢谱、生物合成活性和信号反应。提出了新的 B 细胞分类方案:在四种淋巴组织中鉴定出 12 个独特的亚群,包括 CD45RB + CD27 -早期记忆群体、类别转换的 CD39 +扁桃体常驻群体和有效响应免疫激活的 CD19 hi CD11c +记忆群体。该分类框架和基础数据集为进一步研究人类 B 细胞身份和功能提供了资源。
技术流程
研究结果
1.高度多重的单细胞表面筛选揭示了人类 B 细胞表面蛋白质组
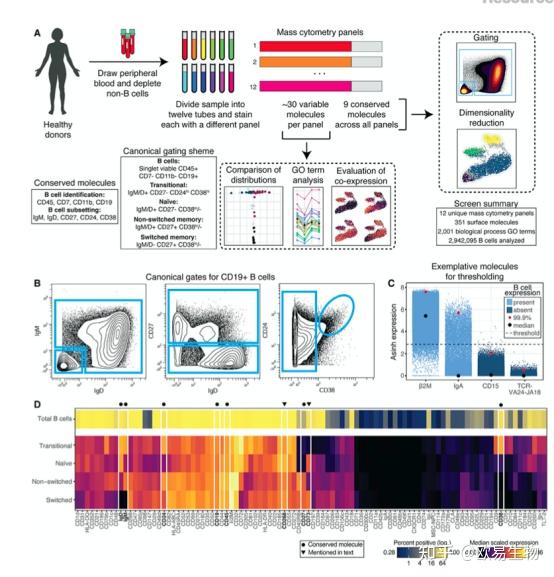
为了识别区分 B 细胞亚群的分子,作者开发了多重筛选的方法,并量化了健康人类 B 细胞上 351 种表面抗原上的共表达模式(图1A)。通过设计了 12 个质谱抗体组,每个组由 9 个用于子集的保守分子和 30 个对每个组独特的可变分子组成。门策略可以实现四个典型 B 细胞亚群:过渡、幼稚、非转换记忆和转换记忆(图1B)。在设置了一个严格的阈值(图 1 C)后,作者确定了 98 个在人类 B 细胞上表达的表面分子(图 1D)。作者的单细胞筛选策略实现了对人类 B 细胞表达的表面分子的可靠鉴定。
图1 |高度多重的单细胞表面筛选揭示了人类 B 细胞表面蛋白质组
a)实验概述 (n = 2 个捐助者)。
b)典型种群的代表性门控。
c)屏幕上分子阳性的代表性阈值。
d)总 B 细胞(顶行)的百分比阳性和 B 细胞表达的分子子集(底行)的中值表达。
2.差异表达分析揭示了幼稚 B 细胞的无反应特性
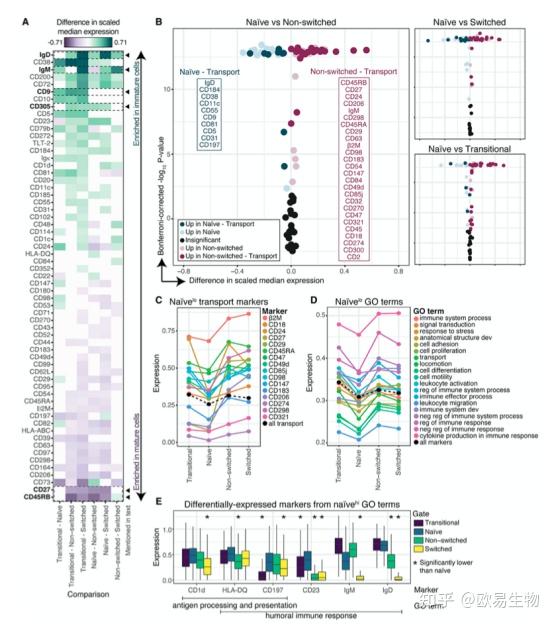
通过规范门控策略识别组织B 细胞的成熟状态:从过渡到幼稚、非切换和切换记忆。为了探究在整个过程中发生的蛋白质组学变化,作者评估了所有分子的子集中每个成对组合之间的表达差异。作者绘制了 61 个差异表达分子(图2A )。
图2 | 差异表达分析揭示幼稚 B 细胞
a)子集的每个成对比较的中值表达差异。所有非白色瓷砖都是显著的(p < 0.005)。
b)比较的火山图,与 GO 术语“运输”相关联。框中列出的显著不同的分子按表达差异幅度的递减排序。
c)转运分子 (颜色) 的中值表达。所有转运分子的中位表达平均值(黑色)。
d)与 GO 术语相关的分子的中值表达平均值 (颜色)。所有分子的中值表达的平均值(黑色)。
e)在幼稚细胞中表达更高的六种分子的表达 (p < 0.005)。
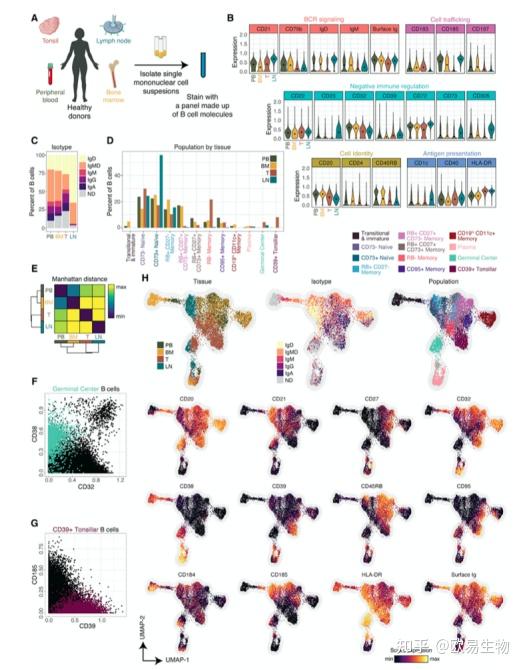
3.CD45RB 标记人类记忆 B 细胞并识别早期记忆群体
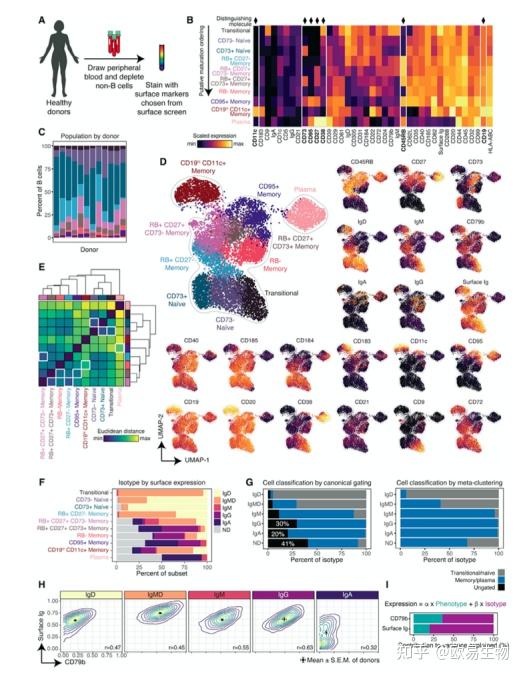
为了找到唯一识别不同 B 细胞的标记,作者以无偏方式分析了所有 B 细胞中分子的共表达模式。作者生成了统一UMAP图,通过使用所有 12 个试管的供体汇集数据来展示保守分子的表达(图 3A)。作者绘制了与保守分子相关的分子,并按功能和相关保守标记进行展示(图 3 B)。
4. 将 B 细胞分为表型和同型不同的亚群
系统筛选了数十种在 B 细胞中差异表达的分子,因此作者假设作者可以将 B 细胞分类为更细粒度的亚群。作者对新鲜、健康的人类外周血 B 细胞(n = 3 名供体)进行了染色,细胞降维成十个不同的群体,包括两个幼稚和六个记忆子集(图 4 B)。表面表达谱提示成熟顺序排列(图4B)。七种不同分子的特征表达以手动门控每个群体,因此也用于标记该方案中的群体:CD11c、CD73、CD95、CD27、CD38、RB 和 CD19(图 4 C D)。子集倾向于在图上形成独特的岛屿,为作者的分类方法提供正交验证 (图 4 D)。
已知 Ig 同种型的使用会影响下游效应器功能和分化模式。因此,作者还在同种型的基础上组织了 B 细胞,并观察到BCR 复合物的两种成分的不同表达模式:表面 Ig 和 CD79b(图4H)。
鉴于这些趋势,作者探索表型或同种型是否对预测表面 Ig 和 CD79b 的表达量贡献更大。作者创建了单细胞多元线性回归模型,其中细胞的表型标记和同型标记用于预测 CD79b 或表面 Ig 的表达(图4H)。尽管两者都提供了丰富的信息,但细胞的同种型对预测两种分子的表达的贡献超过了细胞的表型。总而言之,这些发现表明,作者的高维分类将外周血 B 细胞组织成十个表型不同的亚群,比典型的门控策略更准确地划分细胞。此外,这些表型分区显示出同型限制,这进一步有助于 B 细胞的身份。
图4 | 将 B 细胞分为表型和同型不同的亚群
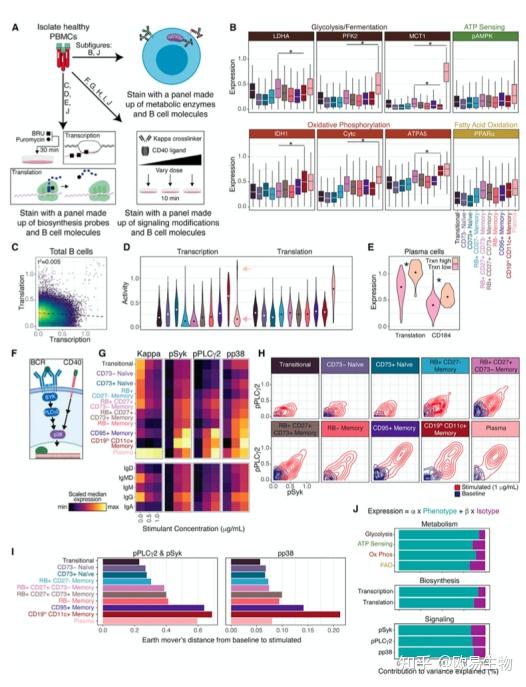
5. B 细胞亚群功能的研究提示了不同的代谢、生物合成和免疫信号活性特征
为了研究作者改进的 B 细胞分类方案的功能特性,作者探索表面蛋白是否表示其他潜在功能细胞过程的差异。作者对来自其他供体(n = 9 个供体)的健康人外周血单核细胞 (PBMC) 进行了染色,并使用质谱仪组来探索 B 细胞代谢谱、生物合成活性和免疫信号传导特征(图 5A)。
研究讨论
为了探究原代细胞的深层表型多样性,作者开发了一种高度多重的单细胞表面筛选,并将其应用于识别可以分离人类 B 细胞亚群的分子。这种方法使作者能够区分四种淋巴组织中的 12 个 B 细胞亚群并关联它们的功能特征。
作者确定了六个记忆群体,证实了先前关于小鼠和人类抗原识别后表型多样化的报道。作者还确定了一个 CD19 hi CD11c +记忆群体,它与在自身免疫、感染和衰老背景下描述的几个群体具有一些共同特征。卡内尔等人,2017)。在这个群体中,作者通过 CD27 表达分离细胞,发现 T-bet 和 PD-1 在 CD27 - CD19 hi CD11c +记忆细胞中富集,类似于在 T 细胞中看到的效应记忆表型。
在这里,作者通过对健康个体中多组学整合进行的深度表型分析揭示了新的、更细的B群体确定,全面映射了人体血液和淋巴组织中的 B 细胞身份。对几个细胞过程中表型与同种型使用的贡献的定量评估突出了对超越谱测序和同种型身份进行分析以了解人类 B 细胞免疫功能的必要性。研究结果作为未来研究在疫苗接种或疾病背景下研究体液免疫反应的资源,描述的群体和分子可能对于理解 B 细胞介导的发病机制或保护至关重要。
小鹿推荐
斯坦福大学研究团队通过大规模流式细胞仪确定了人类 B 细胞表达的 98 个表面分子,确定了四种组织中的十二个B 细胞亚群。
实验技术上,文献采用了质谱流式技术和流式细胞术,以及单细胞代谢谱等先进的单细胞组学技术,形成的框架和基础数据集也构成了研究人类 B 细胞身份和功能的重要资源。

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 






 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 发表于 2025-2-18 15:04
发表于 2025-2-18 15:04