用户名
UID
Email
密码
记住
立即注册
找回密码
只需一步,快速开始
微信扫一扫,快速登录
开启辅助访问
快捷导航
门户
Portal
社区
BBS
资讯
会议
市场
产品
问答
数据
专题
帮助
签到
每日签到
企业联盟
人才基地
独立实验室
产业园区
投资机构
检验科
招标动态
供给发布
同行交流
悬赏任务
共享资源
VIP资源
百科词条
互动话题
导读
动态
广播
淘贴
法规政策
市场营销
创业投资
会议信息
企业新闻
新品介绍
体系交流
注册交流
临床交流
同行交流
技术杂谈
检验杂谈
今日桔说
共享资源
VIP专区
企业联盟
投资机构
产业园区
业务合作
投稿通道
升级会员
联系我们
搜索
搜索
本版
文章
帖子
用户
小桔灯网
»
社区
›
C、IVD技术区
›
化学发光技术
›
【文献翻译】纳米荧光素:一种小荧光素酶点亮了生物发光 ...
图文播报
2026庆【网站十三周
2025庆【网站十二周
2024庆中秋、迎国庆
2024庆【网站十一周
2023庆【网站十周年
2022庆【网站九周年
返回列表
查看:
9863
|
回复:
0
[分享]
【文献翻译】纳米荧光素:一种小荧光素酶点亮了生物发光领域
[复制链接]
wolf
wolf
当前离线
金桔
金币
威望
贡献
回帖
0
精华
在线时间
小时
发表于 2024-12-27 16:59
|
显示全部楼层
|
阅读模式
登陆有奖并可浏览互动!
您需要
登录
才可以下载或查看,没有账号?
立即注册
×
今天给大家翻译的是2016年《
American Chemical Society
》期刊文章:NanoLuc: A Small Luciferase Is Brightening Up the Field of Bioluminescence。
摘要
生物发光蛋白的发现使生物医学领域受益匪浅。目前科学家们将生物发光系统用于许多生物医学应用,从高灵敏度的细胞分析到基于生物发光的分子成像。传统上这些系统是基于Firefly和Renilla荧光素酶;然而这些酶的适用性受到它们的大小、稳定性和发光效率的限制。
NanoLuc(NLuc)是一种新型的生物发光平台,与现有的系统相比具有几个优点,包括增强的稳定性,更小的尺寸,发光增加>150倍。
此外NLuc的衬底显示出更高的稳定性和更低的背景活性,为生物发光成像领域开辟了新的可能性。NLuc系统具有令人难以置信的通用性,可用于广泛的应用程序。NLuc系统的灵敏度提高,稳定性高,体积小,有可能在未来彻底改变报告基因检测领域。然而与所有此类技术一样,NLuc具有局限性(包括体内应用的非理想发射及其独特的底物),这可能导致其在某些分子生物学领域的使用受到限制。随着这项独特技术的不断发展,NLuc可能会在临床前和临床领域产生重大影响,在疾病检测、分子成像和治疗监测方面发挥潜在作用。这篇综述将以一种无偏见的方式向科学界展示NLuc技术,让读者对这个新颖的系统有自己的看法。
1.介绍
自古代文明开始以来,生物发光现象就引起了科学家和普通民众的兴趣。生物发光是发生在某些生物体中产生可见光的化学反应的产物在自然界中,生物体产生生物发光有几个原因包括物种间交流、狩猎和定位食物、吸引猎物和自卫。虽然生物发光已经被观察了几千年,但直到上个世纪分子生物学的进步才使研究生物发光的潜在机制成为可能现在,我们知道
生物发光是一种化学过程,它依赖于酶和底物的相互作用来产生光。所需的酶被称为荧光素酶,底物因荧光素酶的类型(如荧光素)而异。
生物发光反应将在第2节中讨论。
虽然由火和电产生的光会产生大量的热量释放,但“冷光”或由生物发光产生的光效率很高,通过热量产生的能量损失最小。虽然生物医学界已经将生物发光应用于各种应用,但研究人员目前正在开发基于生物发光的产品以帮助一般人群的日常生活。例如利用生物发光的树木和植物来取代传统的路灯的潜力正在被探索。此外一些政府机构已经为研究人员提供了资金以研究军队可能从生物发光中受益的新方法。
虽然一些荧光素酶和相应的底物已经为研究人员提供了十多年,但一些新的荧光素酶直到最近才开始商业化。
NanoLuc荧光素酶(NLuc)是最新的市售荧光素酶。NLuc是一种19.1kDa的荧光素酶,依靠底物呋喃嘧啶产生高强度的发光型发光。
最近NLuc显示出优于传统荧光素酶的几个特性。在这篇综述中我们通过讨论生物发光的最初发现和历史,检查荧光素酶并介绍NLuc系统来研究NLuc如何改变生物医学科学中的生物发光领域。NLuc已用于生物医学研究的多种应用,包括研究蛋白质-蛋白质相互作用、研究遗传调控和细胞信号传导、监测蛋白质稳定性、基于BRET的传感器和分子成像。第3节考察了NLuc在这些应用程序中的适用性并简要讨论了NLuc在这些应用程序中的好处。虽然NLuc被认为优于其他荧光素酶,但第4节研究了NLuc平台与传统荧光素酶系统相比的改进和局限性。最后,第5节总结了主要发现,讨论了NLuc系统的生物医学影响并提供了未来的展望。
2.生物发光技术在生物医学研究中的发展
在过去的十年里随着研究人员有效地利用这种独特现象为生物医学目的带来的好处,生物发光领域得到了极大的发展。不到一个世纪以前第一个荧光素酶被发现。自荧光素酶及其底物的发现以来,生物发光领域取得了重大成就。
2.1.荧光素酶的发现
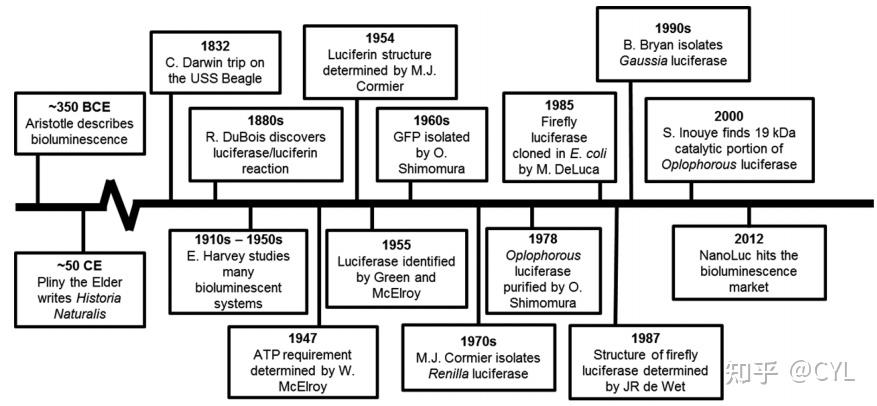
生物发光是一种自然发生的现象,发生在许多陆生和水生生物身上,包括昆虫、细菌、真菌和海洋动物虽然生物发光已经被观察了数千年(图1),但人类直到最近才开始利用这种独特的现象用于生物医学目的。
图1 说明生物发光领域发生的几个重大事件的时间表。
虽然生物发光的第一个参考文献可以追溯到古代文明的民间传说,但历史上第一个关于生物发光的参考文献来自希腊哲学家亚里士多德(公元前384-322年),他对生物如何发光非常着迷。大约三个世纪后老普林尼(公元23-79年)成为第一个报告几种动物生物发光的人,对发光的软体动物、紫色水母、萤火虫和其他动物进行了深入的描述。他的发现发表在一本名为《自然历史》的百科全书中。1667年罗伯特·波义耳发现生物发光需要空气。查尔斯·达尔文也注意到了生物发光,他在贝格尔号上令人难忘的旅行中将这一过程称为磷光。
法国药理学家Raphael Dubois(1849-1929)首先研究了click甲虫生物发光反应所需的成分。拉斐尔·杜波依斯只用冷水和叩头甲虫的腹部就能在实验室里发光。最终他将这两种提取出来的成分命名为“荧光素”,将反应中消耗的分子命名为“荧光素”,将负责反应的酶命名为“荧光素酶”。随着历史的发展,E.Newton Harvey测试了各种荧光素酶和底物的组合,发现荧光素酶和荧光素在物种之间是不可互换的。
虽然杜波依斯在1885年发现了荧光素和荧光素酶之间的反应,但直到1940年代后期Green和McElroy博士才首次提取和纯化萤火虫灯笼。通过这个过程他们分离出酶并确定了它的构象结构。W.McElroy于1947年通过对萤火虫发光的体外研究阐明了萤火虫生物发光所需的ATP。20世纪60年代Osamu Shimomura及其同事从水母
Aequorea victoria
中发现了钙激活的光蛋白水母蛋白。下一个重大发现发生在1985年,当时Marlene DeLuca报道了在大肠杆菌中克隆萤火虫荧光素酶(FLuc),有效地为该技术在未来与许多荧光素酶系统的广泛应用铺平了道路。
在FLuc克隆后的30年里,分子生物学领域发生了一些重要的发现和进步彻底改变了生物发光领域。最终这导致了从
Oplophorus gracilirostris(OLuc)和Gaussia princeps(GLuc)
中发现了荧光素酶和底物。近年来生物发光领域的研究重点是开发高稳定性的荧光素酶。为此一些荧光素酶被发现和修饰,使生物发光在生物医学领域获得了相当大的关注。有关生物发光历史的更多细节,读者可以直接阅读约翰·李的详细综述和参考文献。
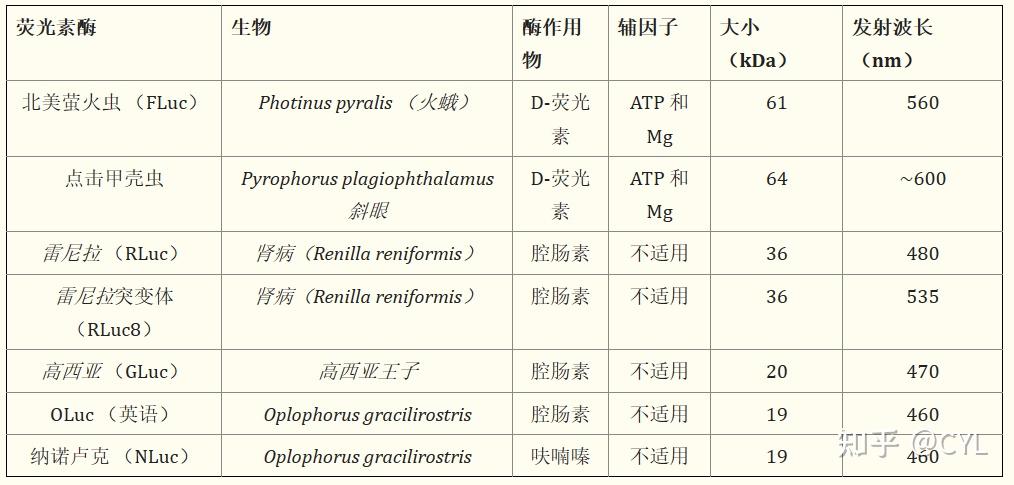
虽然最初认为只有少数生物能够产生生物发光,但进一步的研究表明许多生物都具有这种独特的能力。表1列出了生物医学用途中最常用的荧光素酶。
表1 常见的荧光素酶及其来源、所需的底物和辅因子、分子量和最大发射波长。
最早探索用于生物医学目的的生物发光系统之一是Pyrophorus plagiophthalamus,一种点击甲虫。点击甲虫荧光素酶催化ATP依赖的反应,利用底物D-荧光素以及镁和分子氧使这种荧光素酶类似于其他ATP依赖的荧光素酶如FLuc。点击甲虫荧光素酶的大尺寸可能会极大地影响较小的研究化合物的行为。点击甲虫荧光素酶的衍生物随后被开发出来,具有从绿色到红色的一系列光发射。
20世纪70年代M.J .科尔米耶及其同事从三色堇中纯化出肾形海肾荧光素酶。这种36kDa的酶不依赖于ATP,使用腔肠素和氧气作为辅因子。该系统提供了一种所谓的长闪光发射,一种约30秒的480nm光脉冲。也许最广为人知的用于生物医学目的的荧光素酶是Photinus pyralis或美国萤火虫。J.R.deWet等人于1987年确定了其结构并在哺乳动物细胞中表达了该基因。与海洋生物的荧光素酶不同,这种酶需要ATP和镁以及D-荧光素来产生发光。FLuc和click beetle荧光素酶的结合允许开发双荧光素酶报告物分析,其中可以同时存在实验报告物和内部对照报告物。双荧光素酶系统也与其他平台一起开发包括FLuc和RLuc(可商购)。
最近许多研究人员已经转向更小的荧光素酶。
GLuc是一种荧光素酶,最初提取自海洋桡足类动物Gaussia prince PS。这种荧光素酶与众不同,因为它是一种分泌型荧光素酶
;因此采用这种荧光素酶系统的分析将使用细胞培养基进行分析。与其他海洋生物类似,这种小型荧光素酶(20kDa)产生470nm的光并且不依赖于ATP。布莱恩博士在20世纪90年代分离出这种flash型酶,此后发现它在细胞分泌途径、病毒传染性分析和趋化因子受体激活方面有独特的应用。接下来Shimomura博士于1978年确定了深海细刺裸虾的生物发光特性。
完整的荧光素酶包含两个区域,每个区域由35kDa和19kDa亚单位组成,在其自然状态下使用腔肠素作为底物。然而这种酶已经经历了广泛的发展,19kDa亚基在2000年被确定为生物发光的催化部分。从那时起该亚基已被修改并开发出一种新的底物呋喃嗪以创建称为NLuc的系统。
2.2.荧光素酶:作用机制
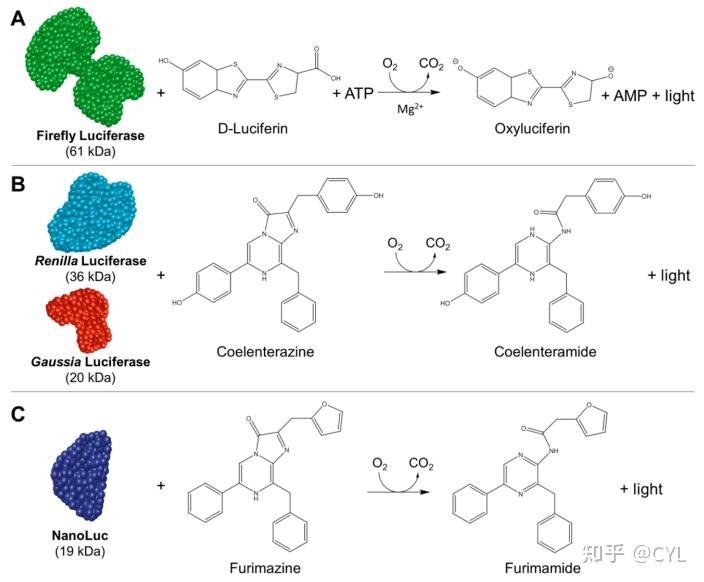
荧光素酶是活生物体中产生光的酶的总称。生物发光的产生有两个关键要求,包括负责催化反应和产生光的酶(荧光素酶)和该酶的底物(荧光素)。许多这样的系统已经被特征化,图2中列出了几个著名的例子。
图2生物发光是基于荧光素酶和相应底物之间发生的化学反应。(A)萤火虫荧光素酶(FLuc)在三磷酸腺苷(ATP)、分子氧和镁存在下与D-荧光素反应产生光。(B)Renilla(RLuc)和Gaussia荧光素酶(GLuc)都只需要腔肠素和氧气来产生光。
(C)当优化的底物呋喃嗪与NLuc在分子氧存在下发生反应时,NanoLuc(NLuc)体系产生生物发光。该反应产生呋喃胺和发光输出。
<hr/>有关荧光素酶机制的详细综述,感兴趣的读者可参考Frank McCapra的精彩解释。简而言之,FLuc与其底物(D-荧光素)之间的反应在存在ATP的情况下产生生物发光(图2A)。接下来RLuc和GLuc与腔肠素反应产生生物发光,而NLuc与腔肠素衍生物呋喃西林反应(图2B,C)。
2.3.纳米Luc系统的开发
NLuc是商业上可用于生物发光应用的荧光素酶系统家族的最新成员。这种小型荧光素酶源自深海虾纤细Oplophorus gracilirostris,在此期间进行了三轮诱变以优化该酶的发光输出。虽然纤细Oplophorus gracilirostris荧光素酶(OLuc)由两个异二聚体区域组成,每个区域具有35kDa和19kDa亚单位,但其生物发光特性已被映射到完整荧光素酶的单个19kDa亚单位,称为
OLuc-19
。虽然该亚单位表达不良且不稳定,但通过诱变和添加新底物对小催化亚单位进行结构优化,发现可以创建非常有效的生物发光系统。此外Mary Hall和她的同事能够优化酶的稳定性。总体而言
他们发现新的NLuc系统显示出比FLuc和RLuc高150倍的比活性。
此外发光输出保持稳定,持续时间比以前的荧光素酶更长。本小节研究NLuc酶及其相应底物的工程设计以改善生物发光。NLuc系统的优势将在第4节中进一步讨论。
虽然NLuc技术相对简单,但为特定应用选择矢量一开始似乎很繁琐。简单回顾一下,表达载体是通过将特定基因引入靶细胞而在细胞中进行蛋白质表达的质粒构建体。有关载体的更多详细信息,请读者参阅基因治疗中该主题的广泛综述。目前有几种NLuc蛋白融合载体可用于产生NLuc与目标蛋白的N-或C-末端融合。它们被分为两种形式,其中pNLF载体系列使用具有多个克隆位点的传统克隆系统,pF系列使用Flexi载体克隆系统,该系统基于使用两种罕见切割限制酶的定向克隆,如果蛋白质编码区在其他Flexi载体之间转移则无需重新测序。
有三种主要形式的NLuc可用于特定应用,包括
非融合形式的NLuc、从细胞中分泌的形式(secNLuc)和称为NLuc-PEST(NLucP)的不稳定形式。
每种形式的NLuc都针对不同的应用进行了配置。非融合形式的NLuc具有最高的灵敏度和光输出,并且由于其细胞内稳定性可以在细胞中积极积累。其次,secNLuc是那些需要将酶分泌到培养基中的应用的绝佳选择。 由于secNLuc在37°C的细胞培养基中保持稳定超过4天。
NLucP是一种独特的蛋白质版本,因为它具有使酶不稳定的蛋白质降解信号从而使NLuc与转录活性和蛋白质表达的变化密切相关,信噪比背景比增加。由于NLucP的寿命较短(10-30分钟),表达水平会随着转录变化而迅速变化。
此外Promega还提供其他基于nluc的技术如NanoBRET,可用于研究蛋白质相互作用动力学以及用于研究信号蛋白的稳定性传感器。有关NanoLuc平台产品的更多信息请访问Promega网站。
3.纳米LUC技术的应用
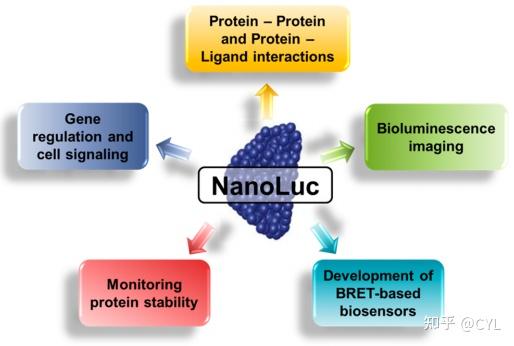
NLuc是一种用途极其广泛的技术,可用于多种应用,包括研究蛋白质和蛋白质配体的相互作用、探索基因调控和细胞信号、监测蛋白质稳定性、开发新型BRET生物传感器和生物发光成像(图3)。
图3 NanoLuc荧光素酶(NLuc)技术已经成功地应用于蛋白质-蛋白质和蛋白质-配体相互作用的研究,基因调控和细胞信号传导的探索,蛋白质稳定性的监测,作为基于bret的生物传感器的利用以及生物发光成像。
本节分析了NLuc在这些应用中的当前用途和优势。
3.1.用于研究蛋白质和蛋白质配体相互作用的NanoLuc
配体受体结合分析是分子生物学的基石。虽然有效,但大多数传统的结合试验依赖于放射性示踪剂的使用,这对使用者构成健康风险,而且许多放射性核素的半衰期很短。为了克服这些限制,科学家们寻找新的非放射性受体结合分析方法,生物发光配体-受体结合分析显示出良好的潜力。特别是生物发光示踪剂显示出比放射性示踪剂更高的灵敏度、更容易制备、更长的保存期和更好的安全性。
开发新型受体结合测定通常既繁琐又耗时,但Song等人开发了一种简化的方法以促红细胞生成素(Epo)为模型快速制备基于NLuc的新型蛋白质示踪剂。糖基化细胞因子Epo与细胞膜受体EpoR结合从而促进红细胞生成形成。在这项研究中NLuc的分泌形式与Epo的C端融合并在人胚胎肾293细胞(HEK-293T)中过表达。Epo-NLuc蛋白保持了对EpoR的高结合亲和力,解离常数(Kd)为0.59±0.06nM,与125I标记的Epo的测量值相似。此外Epo-NLuc的过表达导致大量示踪剂可在转染后4天内获得。使用这些程序有可能在1至2周内制备和测试新的基于NLuc的示踪剂。
NLuc的两个特性使其成为基于细胞的检测的最佳示踪剂,这两个特性是其增强的发光输出和个性化的底物。由于NLuc利用不同于其他荧光素酶的底物,因此有可能通过与其他荧光素酶的多路复用同时研究一种以上的蛋白质-蛋白质相互作用。最近Verhoef等人通过生成可与FLuc复合的分裂NLuc报告子验证了这些特性。
小鼠双微体2(MDM2)同源物和MDM4都是p53肿瘤抑制因子的负调节因子。
因此同时测量这些相互作用可以为复杂的相互作用提供新的见解。NLuc片段与p53和MDM2基因融合。当MDM2和p53相互作用时,这两个NLuc片段会接近从而使该酶重新激活并在存在呋喃西林的情况下产生生物发光。由于NLuc和FLuc使用不同的底物并显示不同的发射光谱,因此有可能研究p53与MDM2和MDM4的相互作用。
在王萍实验室进行的一项研究中使用与琼脂糖珠结合的泛素C端融合的NLuc研究了泛素化过程。泛素化是一种翻译后过程,调节蛋白质降解、蛋白质运输、转录信号和肿瘤发生。在去泛素酶的存在下NLuc从泛素珠释放到上清液中。发光信号与去泛素化酶的活性相关。此外他们将NLuc应用于测量其他泛素样异肽酶和研究OTULIN活性。
2015年Liu等人利用NLuc成功监测了G蛋白偶联松弛素家族肽受体(RXFP3)的内化生成稳定的HEK293T细胞系,共表达Nluc标记的RXFP3和增强型绿色荧光蛋白(EGFP)标记的RXFP3。EGFP用于显微镜观察受体内化过程,NLuc用于生物发光定量测量。由于两种融合蛋白都被编码在一个质粒上,因此使用了一个可诱导的双向启动子来确保两种蛋白的表达是等效的和可控的。正如预期的那样,Nluc标记的RXFP3被内化到酸性溶酶体中导致生物发光信号减弱。此外定量评估了配体诱导的RXFP3内化,暴露于100nM内源性配体松弛素-3的细胞导致受体内化迅速,生物发光信号减弱。在类似的研究中Wu等人研究了人松弛素与RXFP1的相互作用。带着类似的目标,他们用精氨酸取代人松弛素的三个赖氨酸,将NLuc偶联到松弛素上。在毕赤酵母中表达和体外酶促成熟后,松弛素的b链n端与NLuc的c端半胱氨酸之间形成了二硫键。实验构建后,他们发现nluc偶联松弛素对RXFP1保持了较高的亲和力(Kd =1.11±0.08nM),因此该实验适合筛选RXFP1的新型拮抗剂或激动剂。
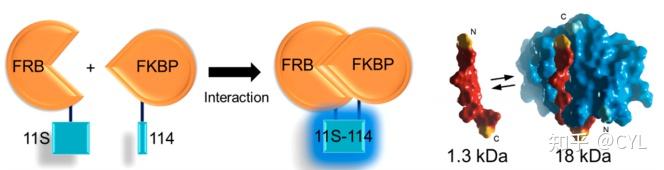
通常使用传统的蛋白质片段互补分析来研究蛋白质相互作用,但这些片段通常未优化且结构受损。Dixon和他的同事们利用NLuc设计了一种新的互补报告基因(NanoBiT)可以对生理条件下蛋白质相互作用进行定量研究。这是通过将NLuc分裂成两种成分来完成的,其中任何一种都将融合到两种感兴趣的蛋白质上。其中一种组分只有11个氨基酸长(114个),而另一种组分的长度是GFP的三分之二(11S)。与其他互补报告不同,NanoBiT在结构上进行了优化以提高构象稳定性并最小化位阻。11S的稳定性得到优化,在生理温度下的表达和性能得到增强,而114的小尺寸保证了它不会影响目标蛋白的相互作用。接下来使用雷帕霉素诱导的FKBP雷帕霉素结合蛋白(FRB)/FK506结合蛋白(FKBP)模型,使用FRB- 11S和FKBP-114融合蛋白确保NanoBiT在哺乳动物细胞中发挥作用(图4)。用等量的NanoBiT或NLuc DNA转染后在活细胞和细胞裂解物中测量发光,在雷帕霉素存在的情况下,在高DNA浓度的NanoBiT和NLuc裂解物中发光呈线性趋势。而在没有雷帕霉素的情况下发光明显减少。
图4利用NanoBiT定量评估生理条件下蛋白质相互作用。利用NanoBiT技术研究雷帕霉素诱导型FKBP雷帕霉素结合蛋白(FRB)/FK506结合蛋白(FKBP)在哺乳动物细胞中的相互作用,采用FRB- 11s和FKBP-114融合蛋白。经参考文献28许可改编。版权所有2015年美国化学会。
两项研究利用NLuc系统研究了
铁外排转运蛋白即人铁转运蛋白(Fpn)
。首先Song等人将NLuc连接到Fpn的C端以监测该受体在哺乳动物细胞中的内化。此外这种融合蛋白用于定量评估Fpn与肽激素(称为铁调素)的相互作用。在hepcidin处理和不处理的情况下,对表达FpnNLuc的完整细胞和裂解细胞进行发光测量。hepcidin处理后裂解细胞的生物发光信号略有下降,这归因于NLuc的高稳定性。然而hepcidin处理后完整细胞的发光明显减少50%,这与溶酶体中的低pH值有关。通过测量pH值为4−5时NLuc的活性证实了这一点,pH值与溶酶体的酸度相似。接下来同一课题组进一步利用NLuc研究胰岛素样肽3(INSL3),其与松弛素家族肽受体2(RXFP2)的相互作用在生育中发挥了重要作用。在这项工作中作者建立了一个简单的程序,通过首先创建一个完全活跃的
6×Histidine-cysteine-NLuc(6×His-Cys-NLuc)
可以与巯基蛋白残基反应将NLuc与感兴趣的蛋白质化学连接起来。同样,后来He等人利用6×His-Cys-NLuc研究了人白血病抑制因子(LIF)与其天然受体(LIFR)的结合。正如预期的那样,LIF-NLuc显示出不变的结合亲和力,每个细胞的检测限小于10个受体,使该平台成为超灵敏的生物发光示踪剂。
在一个类似的研究中NLuc被用于研究体外肌醇要求酶1(IRE1)的活性IRE1的激活是由内质网(ER)的异常状况引起的,这与几种神经退行性疾病有关。当这种情况发生时未剪接形式的X-box结合蛋白1被切割产生sXBP1。以小鼠XBP1splice区域为上游设计了NLuc报告结构;因此不同内质网应激诱导剂可诱导XBP1-NLuc的表达。为了评估报告蛋白在体外的效率将XBP1-NLuc转染到4个细胞系中,然后用内质网应激诱导剂处理。结果表明暴露于内质网应激诱导剂后NLuc活性增加。在同一组的另一项研究中NLuc被用于监测HEK293细胞中脑星形细胞源性神经营养因子(MANF)的转录调节。HEK293细胞分泌Nluc标记的MANF具有时间依赖性并且可以被两种已知的减少野生型MANF分泌的蛋白减慢。此外刺激细胞产生更多的MANF,这导致细胞培养基中测量的NLuc活性显著增加。该方法可应用于研究其他内质网应激性疾病,包括肌萎缩侧索硬化症、缺血和糖尿病。
NLuc的潜在应用超出了内源性蛋白质的研究,包括病毒感染的研究。最近Seay等人利用NLuc构建了人类免疫缺陷病毒(HIV)报告结构。虽然RLuc最初是为这项研究开发的,但作者希望提高该检测的灵敏度和稳定性。NLuc增强的灵敏度对于这种应用很重要,因为它允许在初始病毒感染期间定量低水平HIV复制。由于研究人员想要研究急性HIV感染和暴露前预防(PrEP)通过阴道粘膜抑制HIV感染的功效,因此使用了来自最近感染HIV的非洲妇女的R5-tropic env基因(C.Du151.2)。在293T细胞中产生病毒后他们在细胞中发现了高水平的NLuc活性,这表明NLuc的表达稳定且高度敏感。此外他们还能够监测病毒感染的初始阶段。此外该小组后来开发了一种名为hCD4/R5/cT1的新型小鼠菌株,这是一种携带HIV-1感染必需基因的转基因菌株,包括人分化簇4(CD4),C−C趋化因子受体5型(CCR5)和细胞周期蛋白T1。该小鼠模型感染了表达HIV-1的NLuc,这使得研究药物滥用和其他因素如何通过粘膜传播增加HIV-1感染的风险成为可能。
NLuc也被用于研究其他病毒感染,包括乙型肝炎病毒(HBV)。Nishitsuji等人开发了一种名为HBV/NLuc的重组HBV,用于监测病毒在体外的复制。这是通过在HBV控制区下游插入NLuc基因来实现的。必须保持HBV DNA大小不变,因为如果基因组大小超过原始基因组大小0.7千碱基对(kbp),复制就会受到严重阻碍。结果表明NLuc活性准确地反映了转染细胞中的HBV RNA水平,因为在HBV感染后6天以上两者都增加了。接下来新的HBV/NLuc报告基因被用于筛选HBV抑制剂,包括恩替卡韦和干扰素-β(IFN-β),这两种抑制剂都会导致NLuc活性的剂量依赖性降低。虽然qPCR和qRT-PCR是评估HBV感染的常用方法,但HBV/NLuc系统被证明更简单、更敏感。该检测利用NLuc监测病毒复制的独特特性,可能在未来用于鉴定新的抗hbv药物和抗体。
除病毒外NLuc的应用还扩展到其他病原体,如引起疟疾的寄生虫恶性疟原虫。虽然治疗和患者控制方面的进步有效地减少了疟疾病例的数量,但据世界卫生组织统计,每年仍有40多万人死于疟疾。为了开发一种新的高通量筛选方法来筛选抗疟疾化合物,Azevedo等人描述了一种在恶性疟原虫中表达NLuc的方法。作为概念验证,表达NLuc的寄生虫被证明可作为测定抗疟化合物抑制剂效果的功能性筛选试验。类似的方法也被用于在细菌中表达NLuc。在Thomas Proft博士实验室进行的一项研究中NLuc被用于产生生物发光的A型链球菌(GAS)。菌株结果表明Nluc修饰的细菌比FLuc修饰的GAS亮15倍以上。然而NLuc-GAS不能用于区分代谢活跃和不活跃的细菌,因为NLuc不依赖于ATP。因此NLuc在未来的潜在应用中可能会受到限制,在这种应用中ATP依赖的FLuc将发挥更好的作用。
3.2.探索基因调控和细胞信号的纳米技术
基因递送是分子生物学领域的一个重大进步,分子生物学在生物医学界迅速受到关注。为了确保基因传递的成功可以使用某些标记。在最近的一项研究中NLuc被用于证实在小鼠中通过水力肢体静脉注射(在尾静脉中快速注射大量质粒DNA)成功递送线粒体基因而无需使用病毒进行转染。使用内源性线粒体蛋白(称为人NADHubiquinone氧化还原酶亚单位4(ND4))的DNA设计了一种基因构建体。作者预计从ND4-NLuc载体转录的mRNA将有效地与线粒体核糖体结合以翻译成可检测的蛋白质。虽然蛋白质印迹法缺乏检测线粒体转染外源蛋白的灵敏度,但NLuc允许使用这种新型结构验证肝脏和骨骼肌中的基因递送。
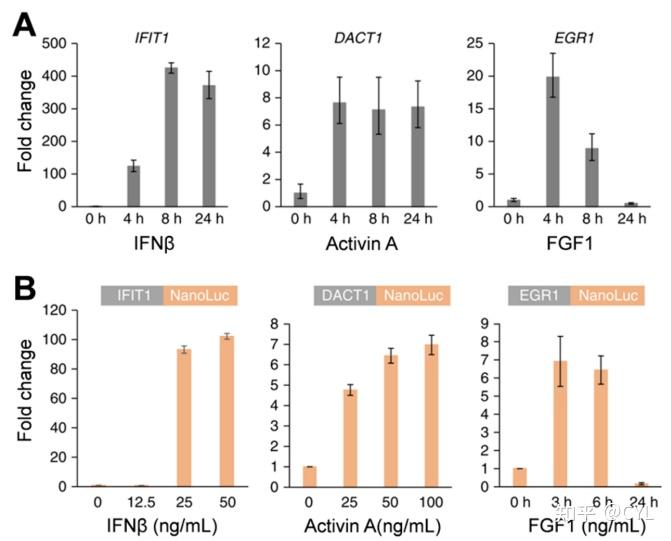
Lackner等人还利用NLuc的小尺寸和亮度选择了一种与单次基因组整合事件产生的低信号强度兼容的系统。该研究检查了一种在N端和C端标记基因的新策略,使用NLuc来演示该方法。NLuc标记的细胞系用于监测内源基因表达。该细胞系在三种细胞因子诱导基因(DACT1、IFIT1和EGR1)中含有NLuc整合。首先使用定量PCR在野生型HAP1细胞中评估mRNA对细胞因子刺激的上调(图5A)。为了测试NLuc的功能,用同源细胞因子配体(对每个基因特异)刺激DACT1、IFIT1和EGR1中携带NLuc整合的细胞系,这导致细胞因子诱导的NLuc上调(图5B)。重要的是表明NLuc关于细胞因子刺激和动力学的数据与qPCR数据具有良好的相关性。此外标记的等位基因在细胞定位和基因表达调控方面仍保持完全功能。最终NLuc报告细胞系被证明可以在活细胞的内源水平上监测基因表达。
图5利用细胞系表达NanoLuc(NLuc)整合物监测基因表达。(A)用IFN-β、激活素A和FGF1三种细胞因子刺激HAP1细胞4、8或24小时,最终浓度为50ng/mL。分离RNA并分析IFIT1、DACT1和EGR1的特征。
(B)三种细胞因子刺激具有NLuc整合IFIT1、DACT1和EGR1的克隆细胞系。在收集细胞系的IFIT1和DACT1的24h以及EGR1的记录时间点之后测量NLuc水平。转载许可来自参考文献41。自然出版集团版权所有。
<hr/>监测基因表达对于理解从病毒感染到基因传递的各种现象至关重要。除了研究基因表达外,新型遗传报告基因的发展使得研究与基因表达相关的细胞事件成为可能。生物发光报告子已被用于研究RNA干扰(RNAi)、各种启动子的结构以及启动子单核苷酸多态性(SNPs)的功能分析;因此我们预测未来的研究将采用NLuc进行类似的应用。
3.3.用于监测蛋白质稳定性的NanoLuc
蛋白质聚集已被发现是几种人类疾病的关键因素,从亨廷顿舞蹈症到阿尔茨海默病。虽然蛋白质聚集的作用已经得到了很好的证明,但在体外和体内研究蛋白质聚集的方法有限。为了成功地研究蛋白质聚集并促进针对蛋白质聚集的新治疗策略的发展,需要高度敏感的药物。NLuc结构是探索这种生物学现象的可能平台,展示了可能受益于这项新技术的另一种生物医学应用。
考虑到这些限制,Zhao等人使用条件分裂-NLuc系统监测活细胞中的蛋白质聚集。NLuc的一部分(N65)融合到已知聚集的蛋白质上,而NLuc的另一部分用于测量蛋白质聚集的水平(图6A)。
图6 NanoLuc(NLuc)片段用于测定蛋白质聚集。(A)聚集实验示意图,其中蛋白质与NLuc的N端融合用于监测蛋白质聚集,发光信号减少表示蛋白质不溶性。(B)添加呋喃马嗪50min后单个NLuc片段和组合片段的发光输出。
(C)作为概念验证,单个或两个NLuc 片段在哺乳动物细胞中表达。发光需要NLuc片段N65和66C的存在。经参考文献42许可改编。版权所有2016年美国化学会。
<hr/>如果蛋白质-NL融合聚集,则添加第二部分NLuc在添加底物后不会产生生物发光。如果蛋白质没有聚集,加入NLuc的第二部分在加入呋喃嘧啶后会产生生物发光信号。在体外对NLuc的两个片段进行了表征以确保这两个片段(N65和66C)都是产生发光信号所必需的(图6B)。最后,作者将这种技术从细菌转移到哺乳动物细胞作为原理证明,表明只有当NIH-3T3细胞共转染表达两个NLuc片段时才会观察到发光(图6C),进一步验证了NLuc在体外监测蛋白质聚集的用途。
3.4.基于BRET生物传感器的NanoLuc
生物发光共振能量转移(BRET)使得在生理温度下实时研究活细胞中的分子相互作用成为可能。在BRET中荧光素酶作为共振能量的供体,而荧光蛋白是共振能量的受体。在添加荧光素酶底物后,来自荧光素酶的生物发光能量激发荧光团,避免了对激发光的需要并防止了荧光团可能的光漂白。优化的BRET平台利用具有高生物发光量子产率和优异光谱分辨率的供体。此外由于组织衰减,BRET成功转化为小动物成像需要红移信号。例如之前使用突变的RLuc8(λex/em=480nm,coelenterazine)和mOrange(λex/em=514/530nm),因为这种BRET反应将能量转移到530nm。
RLuc是BRET研究中常用的能量供体,与黄色荧光蛋白(YFP)匹配。该方法虽然有效但受背景噪声大、灵敏度低的限制。与RLuc相比,NLuc的生物发光光谱窄得多,更容易区分受体荧光团。另一种称为HaloTag融合标签的新技术允许对几种潜在的荧光团进行快速评估。
HaloTag系统是基于一个稳定的融合蛋白的形成,该融合蛋白携带一个用于结合应用特异性配体的活性位点。
这导致了HaloTag标记的蛋白和特定配体之间的共价相互作用。这两种技术都被用来研究雷帕霉素诱导的FKBP12与Frb之间相互作用的BRET效率,因为这种简单的模型通常用于研究新的蛋白质-蛋白质相互作用。首先确定了最优的HaloTag配体为非氯tom(NCT),其激发和发射最大值在595/635nm处,与NLuc相比红移超过175nm(图7A)。为了比较我们还研究了RLuc和YFP之间的BRET效率(图7B)。创建两个构建体并将基因转染到细胞中(图7C)。接下来将细胞与NCT孵育,NCT与HaloTag修饰的蛋白形成共价键。然后用呋喃嘧啶孵育细胞,呋喃嘧啶与NLuc反应产生发光,能量有效地转移到NCT。这验证了基于Nluc的BRET在活细胞中监测蛋白质相互作用的潜力。
图7 NanoBRET用于检测Frb和FKBP之间的蛋白-蛋白相互作用。
(A)在NanoBRET中NLuc与NCT之间存在175nm的差异。
(B)以RLuc8和YFP作为对照系统的BRET,信号距离为44nm。
(C)NanoBRET系统示意图,其中两种蛋白与NLuc或HaloTag融合并在哺乳动物细胞中表达,用于体外BRET成像。经许可改编自ref47。版权所有2015年美国化学会。
<hr/>最近Robers和他的同事开发了一种方法,该方法利用一种细胞渗透性荧光蛋白,结合活细胞中的NLuc融合细胞内靶蛋白。使用与靶蛋白结合的化合物,表明BRET信号与相互作用分子的结合效率相关。因此相互作用分子与靶蛋白的结合将导致可用于结合荧光示踪剂的靶蛋白减少从而导致BRET信号减弱。Schaub等人的一项类似研究开发了一种用于体内使用的新型BRET成像报告分子,称为LumiFluors。构建了两种LumiFluors,即eGFP-NLuc (GpNLuc)和LSSmOrange-nano Luc(OgNLuc),它们通过荧光团C端和NLuc N端之间的柔性肽接头连接在一起。从这两种发光物质获得的分子内BRET信号在注射了呋喃西林的A549(非小细胞肺癌)皮下和原位小鼠中产生了迄今为止已知的最亮的生物发光信号。此外,这种新型系统对检测局部淋巴结中的微转移足够敏感。
NLuc也被用于研究胰岛素与胰岛素受体的结合。胰岛素受体的两个胰岛素相互作用结构域(αCT和L1结构域)分别与NLuc或荧光蛋白YPet进行基因融合。结合胰岛素后,这两个胰岛素相互作用结构域相互靠近,有效地使NLuc和YPet紧密接触以产生BRET信号。发现BRET信号与样品中胰岛素的量成比例。NLuc在BRET的使用使研究人员能够开发高度敏感的结构,用于可视化从胰岛素结合到癌症检测的过程。虽然体外测试是生物医学领域的一个重要工具,但临床前转化需要在动物模型中进行进一步研究。
3.5.用于体内生物发光成像的NanoLuc
在过去的十年中,生物发光已经成为体内光学成像的一种广泛使用的方法,因为生物发光技术允许低背景噪声和高灵敏度的非侵入性分子成像。由于这些原因,生物发光成像在生物医学研究的临床前环境中越来越受欢迎。
传统上生物发光成像是通过FLuc进行的,因为其560nm的强发射可以对几厘米深的组织进行成像。迄今为止其他传统荧光素酶在体内的应用仍然受到限制,因为它们的光谱发射谱对于深度组织成像不是最佳的。
虽然NLuc没有用于体内成像的最佳光谱发射,但它已被用于多种成像应用,包括追踪病毒感染和监测疾病进展。
在Stacer等人的研究中将NLuc用于乳腺癌双光学成像,方法是将分泌NLuc或Gluc的质粒瞬时转染MDA-MB-231人乳腺癌细胞将转染Nluc的细胞植入小鼠乳腺脂肪垫。小鼠注射呋喃嗪后成像显示无滤光片信号最高,其次是500 nm滤光片(图8A)。接下来将转染NLuc和葡萄糖的肿瘤异种移植物植入小鼠体内以比较两种生物发光剂之间的差异。注射5μg furimazine可提供来自MDAMB-231-NLuc肿瘤的强烈光信号,而来自MDA-MB231-GLuc肿瘤的信号保持在背景水平(图8B),进一步证明了furimazine对NLuc的底物特异性并表明该双成像系统具有巧合报道的潜力。
图8 NLuc与GLuc在乳腺癌影像学中的比较。用NLuc或GLuc转染MDA-MB-231细胞。(A)转染NLuc的MDA-MB-231细胞植入乳腺脂肪垫小鼠的生物发光成像。注射呋喃嘧啶后分别用开放滤光片、500nm滤光片和520nm滤光片获得图像。
(B)植入NLuc或表达glu的MDA-MB-231肿瘤异种移植物的小鼠注射coelenterazine并成像2分钟。SAGE出版社版权所有。
<hr/>此外NLuc在每个成像时间点都比GLuc显示出增强的信号。这也证实了NLuc可以用于浅表肿瘤和内脏器官的体内成像。为了确定多路复用能力,还使用MDA-MB-231-NLuc/FLuc细胞同时表达NLuc和FLuc来监测肿瘤进展。虽然这种生物发光系统能够有效地监测肿瘤生长,但NLuc的发射波长(~460nm)比发射波长大于600nm的红移FLuc更不适合用于深部组织成像。
Germain-Genevois等人也研究了NLuc与FLuc的双重报告者成像,特别是在深部脑肿瘤和全身转移中。U87-FRT-CMV-NL-IRES-FL肿瘤使用NLuc(furimazine), FLuc(D-荧光素)底物和coelenterazine(一种适合这两种底物的底物)进行成像。使用coelenterazine产生来自两种荧光素酶的信号,表明即使在同一肿瘤内单次底物注射也可以连续测定NLuc和FLuc的活性。NLuc在体外表现出更高的活性,而FLuc在体内表现出更强的活性,因为它更适合于组织穿透发射。在一个类似的研究中将表达NLuc或FLuc的病毒载体注射到小鼠的脚垫中并成像以跟踪病毒的传播。在个别阳性组织中FLuc的信号明显增加,但注射后48小时内NLuc的全身信号持续存在,比同一时间点的全身FLuc信号高约2个log。由于NLuc和GLuc在coelenterazine存在下都会产生光,因此NLuc和GLuc双报告基因成像确实需要一些优化。幸运的是Heise等人解决了这一问题,他们表明可以通过测定NLuc在修饰底物furimazine存在下的活性来计算含有coelenterazine的NLuc的背景信号。反过来,GLuc的活性可以计算,因为它不与呋喃嘧啶反应。这一发现为高灵敏度双荧光素酶系统开辟了新的途径,它可能对高通量筛选有用。
长期以来,生物发光报告基因一直被用于动物传播病毒的成像。然而包括流感在内的许多病毒病原体对其基因组的变化特别敏感,因此无法插入大型传统报告基因。NLuc具有19kDa的小尺寸克服了这一障碍并通过生成编码PA-NLuc融合的报告病毒,首次在小鼠模型中对流感传播进行体内跟踪NLuc病毒产生的通量。随着小鼠体重减轻和感染变得更严重而增加。由于雪貂被普遍认为是最接近人类免疫系统的模型,Karlsson等人也证明了NLuc在该模型中成像流感传播的能力来自鼻腔通道的生物发光与病毒滴度显著相关并且在感染雪貂的所有扫描中都很容易看到。呼吸道感染则不太容易被发现,由于覆盖组织的存在肺部的流量减少了高达4.5倍。
4.纳米LUC技术的改进和局限性
几乎所有讨论的应用都可以使用其他荧光素酶系统,那么NLuc系统与传统荧光素酶系统相比有什么优势呢?
NLuc平台的优势包括其增强的亮度、热稳定性、pH稳定性以及在细胞中的无偏分布。此外与绿色荧光蛋白(GFP)等其他报告基因不同,NLuc不需要在哺乳动物细胞中进行翻译后修饰。
这些优势使得在更具生理相关性的条件下研究生物学成为可能,在这些条件下报告基因可以低水平表达。此外NLuc在多种应用中显示出特殊优势。更具体地说,NLuc亮度的提高和分析活细胞的能力使得使用发光显微镜进行细胞成像更加可行。虽然该领域一直由荧光蛋白共聚焦显微镜(xFP)占据主导地位,但NanoLuc允许进行出色的宏观检测和细胞成像以确认和研究生物学。此外NLuc系统对以前的类似技术进行了改进,因为NanoBiT和NanoBRET可用于体外和体内检测,可以可逆地测量蛋白质的相互作用。由于这些系统是可逆的,因此它们也可用于监测与竞争性抑制剂一起培养时相互作用蛋白质的位移。
当考虑目前市场上可用的荧光素酶平台时GLuc与NLuc具有最多的相似性。回顾一下,这两种荧光素酶系统和其他系统(例如RLuc)都来自海洋物种;因此这些酶通常比来自其他生物的荧光素酶小(表1)。此外,来源于海洋动物的荧光素酶不依赖于ATP并利用腔肠素作为其底物,NLuc除外。几项研究表明与荧光素相比,腔肠素稳定性差、溶解性差、毒性大且价格昂贵。另外GLuc是一种分泌型荧光素酶,因此它在潜在应用中受到限制;而非分泌型和分泌型NLuc平台均已建立。根据Hall等人的报告,GLuc的光强会迅速衰减。在大多数情况下GLuc反应产生的光会迅速衰减,需要配备注射器的照度计进行测量。NLuc可以用PEST降解信号标记,这允许荧光素酶表达的快速变化,这种变化在对转录动力学的响应中更持久。虽然目前没有GLuc-PEST的报道,但GLuc和其他荧光素酶可以以类似的方式进行调节。随着科学家寻找新的遗传操作的荧光素酶系统,已经进行了大量的努力来改善这些局限性。
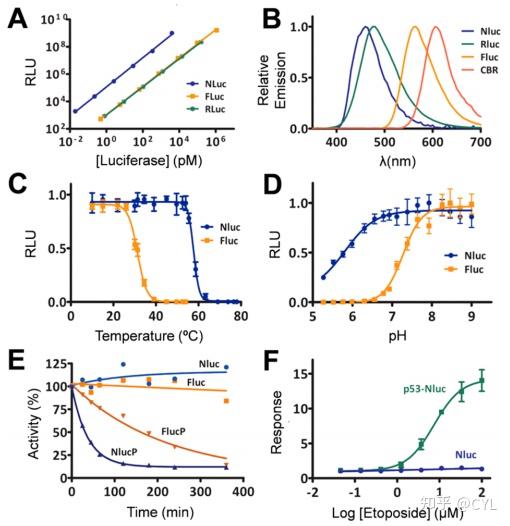
开发新的荧光素酶系统不是一项简单的任务,因为最佳情况要求酶是小的单体并且在酸性/碱性条件下和高/低温下高度稳定。正是对OLuc-19的基因操作导致了NLuc系统的形成。结合酶特异性底物呋咱,这项新技术超出了大多数预期,因为它为众多生物发光应用提供了一个高度通用的荧光素酶平台。在NLuc的合成过程中进行了几项研究以比较几种荧光素酶的生物发光亮度(
图9A
)、最长发射(
图9B
)、温度稳定性(
图9C
)、pH稳定性(
图9D)
。此外,还测定了NLucP的不稳定特性并与FLuc的不稳定形式进行了比较,显示出更快的响应速度(图9E)。最后,表达p53-NLuc的细胞表明在用依托泊苷处理后可以监测细胞中p53的积累,增加依托泊苷浓度导致更高的信号(图9F)。
图9 NLuc、FLuc和RLuc在10min时的发光强度比较。NLuc、RLuc、FLuc和click beetle red luciferase(CBR)的光谱分布,显示出较宽的发射峰范围。比较NLuc和FLuc对(C)温度和(D)pH的敏感性。(E)环己亚胺抑制蛋白合成后随时间的发光测量结果表明NLuc-PEST(NLucP)信号迅速下降,其次是FLucP。
(F)NLuc通过在HEK293细胞中表达p53−NLuc来研究p53稳定性的调控变化。依托泊苷的加入引起了p53的积累,这可以用呋喃嘧啶来测量。经参考文献4许可改编。版权所有2012年美国化学学会。
<hr/>虽然NLuc与传统的荧光素酶系统相比有几个优点,但该系统有一些限制,可能需要解决这些限制以进一步实施该技术。NLuc平台的第一个限制是其发射光谱,这对于体内研究来说不是最佳的。NLuc的光谱分布最大发射波长为 460nm,相对于RLuc和GLuc分别约为20和25nm蓝移。几个荧光素酶平台已经经过遗传操作将其最大发射光谱转移到红色区域,包括Rluc8(520nm)和另一种被称为Green-RLuc(560nm)的遗传修饰的RLuc。此外click beetle荧光素酶平台以其610nm的红移光谱发射而闻名。通过将这些荧光素酶系统转移到红色光谱区,它们的潜在应用随着体内使用变得可行而拓宽。虽然对于在NIR光谱区工作最好的体内成像来说仍不是最佳的,但这些是生物发光成像领域的重大改进。由于NLuc不是体内成像的最佳候选,第3节中描述的几项研究利用了NLuc的BRET功能来有效克服这一限制。应该考虑的一个潜在好处是NLuc平台与其他荧光素酶系统的复用能力,允许双荧光素酶分析具有充分分离的光谱发射谱以获得更大的动态范围和更高的灵敏度。
NLuc平台的另一个潜在限制是对呋喹嗪的要求,呋喹嗪是一种仅适用于NLuc的底物,一般无法获得。因此NLuc平台的成本高于传统的荧光素酶系统,这可能会有效地限制其在许多实验室的可用性。随着该系统的完善和在科学界的广泛应用,我们相信成本将不再是一个障碍。考虑到这些概念,NLuc在生物医学领域的应用不断增长。例如最近在Nature Methods上发表了一份协议,描述了如何使用NLuc平台监测配体与GPCRs的结合。在该协议中作者提供了NLuc和RLuc8之间的直接比较并表明NLuc在哺乳动物细胞中产生的生物发光信号比RLuc8大得多。此外NLuc对受体的正常活性或细胞追踪没有表现出不利影响而RLuc8则导致受体运输的改变。荧光素酶系统之间的直接比较是非常宝贵的,因为它们为研究人员提供了做出哪种荧光素酶系统最适合特定应用的明智决定所需的事实。
5.结论和未来展望
随着新型荧光素酶的开发,生物发光领域不断发展。由于NLuc是荧光素酶家族的最新成员,研究人员仍在研究该系统的潜在用途。这篇综述中讨论了NLuc技术的几种应用,包括蛋白质-蛋白质和蛋白质-配体相互作用的研究。尽管一些研究已经将NLuc用于该应用,但仍需要新的高通量筛选分析。尽管NLuc在这方面表现出了前景,但仍然很难监测体外或体内发生的一种以上的相互作用。由于细胞中每分钟都有数千种相互作用发生,新型生物发光多重技术的发展将大大提高我们对疾病进展和治疗干预的理解。
NLuc也被证明是监测细胞信号和基因调控的有效工具。由于基因递送是分子生物学家的中心主题,NLuc系统可以进一步扩展以监测体内基因递送的成功。
虽然NLuc目前受到其蓝色光谱发射的限制,但科学家可能能够在未来通过遗传操作这种酶来产生远红移的NLuc从而保持其增强的稳定性和卓越的亮度。
接下来使用NLuc监测蛋白质稳定性;然而NLuc尚未针对此应用进行广泛调查。由于几种疾病与蛋白质聚集直接相关,如阿尔茨海默氏症,NLuc系统可能有望在未来检测新的蛋白质不稳定途径。NLuc的第四个应用是用作BRET生物传感器。
除了这里提到的应用之外,NLuc的多功能性超越了哺乳动物。几项研究利用NLuc监测病毒和细菌。由于生物发光分析相对快速且廉价,未来的研究可能会采用基于NLuc的分析来检测或监测患者样本中的某些感染。此外NLuc的亮度增强使早期检测疾病变得可行,包括艾滋病毒或一些细菌感染。
由于NLuc技术仍是新技术,许多研究人员对从传统荧光素酶系统转换仍持谨慎态度。在某些情况下NLuc可能不是特定应用程序的最佳选择。然而对于那些由于生物发光信号低而停止研究的研究人员来说,NLuc可能是一个很好的选择。随着生物发光领域的不断发展,新型荧光素酶的开发将继续扩大可能性的范围。
<hr/>
PDF原文
下载链接:
原文地址:https://zhuanlan.zhihu.com/p/685306516
回复
举报
返回列表
发表回复
高级模式
B
Color
Image
Link
Quote
Code
Smilies
您需要登录后才可以回帖
登录
|
立即注册
本版积分规则
发表回复
回帖后跳转到最后一页
浏览过的版块
PCR技术
同行交流
病理检验
科技前沿
行业研究
关闭
官方推荐
/3
AI助手<小桔子>来了!
欢迎来交流,可以回答IVD行业各类问题!
查看 »
IVD业界薪资调查(月薪/税前)
长期活动,投票后可见结果!看看咱们这个行业个人的前景如何。请热爱行业的桔友们积极参与!
查看 »
小桔灯网视频号开通了!
扫描二维码,关注视频号!
查看 »
返回顶部
快速回复
返回列表
客服中心
搜索
洽谈合作
关注微信
微信扫一扫关注本站公众号
个人中心
个人中心
登录或注册
业务合作
-
投稿通道
-
友链申请
-
手机版
-
联系我们
-
免责声明
-
返回首页
Copyright © 2008-2024
小桔灯网
(https://www.iivd.net) 版权所有 All Rights Reserved.
免责声明: 本网不承担任何由内容提供商提供的信息所引起的争议和法律责任。
Powered by
Discuz!
X5.0 技术支持:
宇翼科技
浙ICP备18026348号-2
浙公网安备33010802005999号
快速回复
返回顶部
返回列表

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 






 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 发表于 2024-12-27 16:59
发表于 2024-12-27 16:59