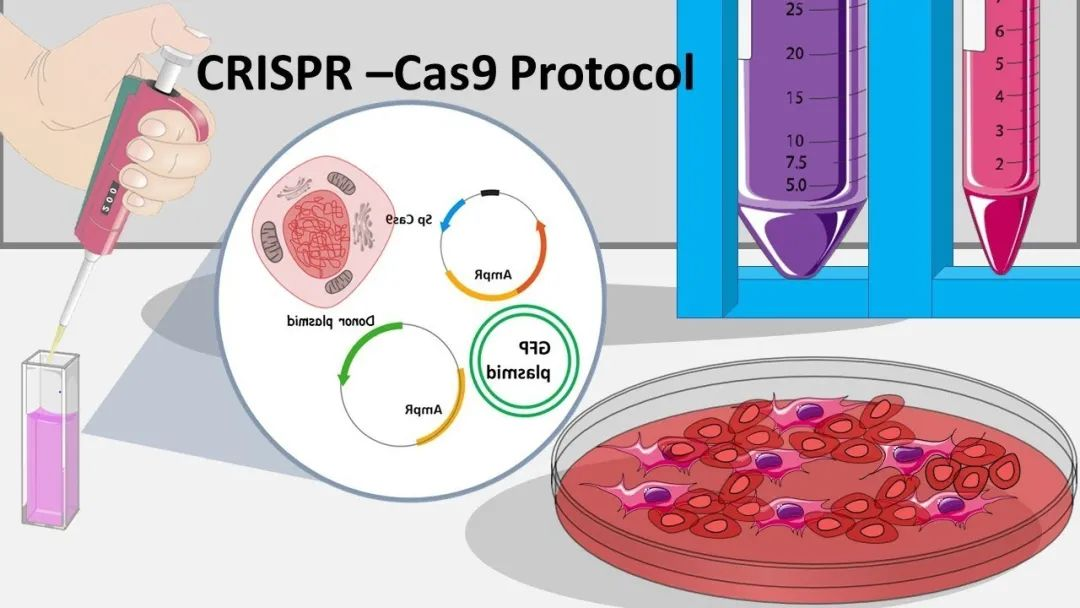
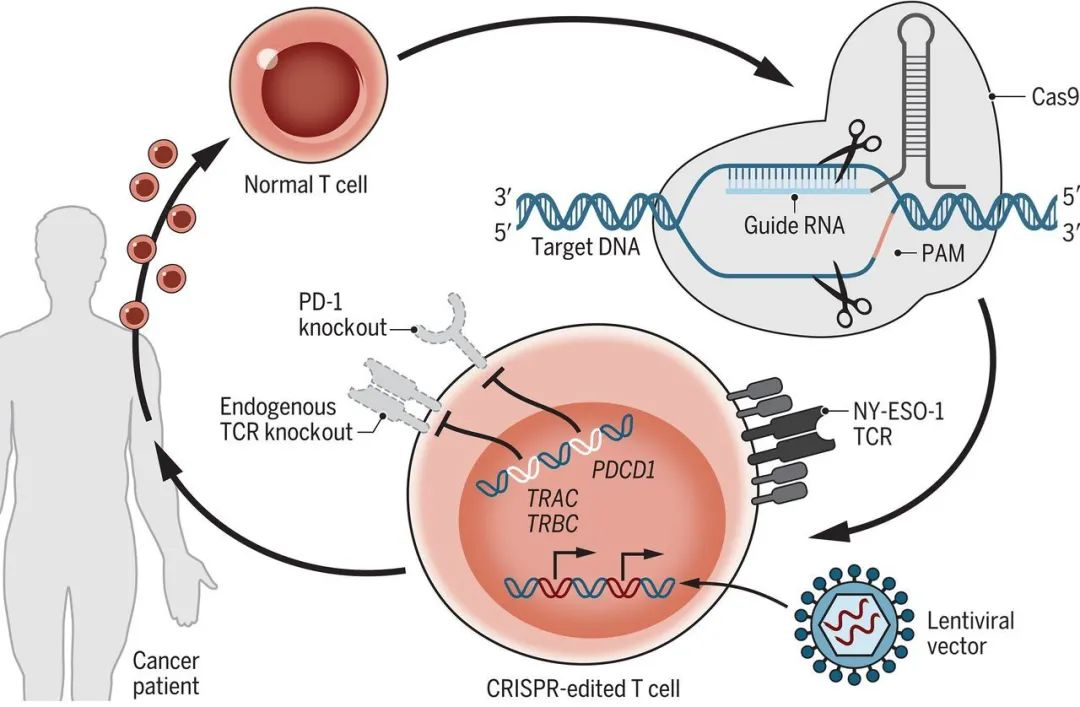
下面简单介绍下CRISPR这个神仙技术的简史。
CRISPR的发现及其功能
1993年报道,弗朗西斯科·莫吉卡(Francisco Mojica)是第一个描述现所谓的CRISPR基因座的研究者。2000年,他发现完全不同的重复序列其实是具有一组共同的特征,即现在已知的CRISPR序列的特征,在2002年首次在印刷品中使用了CRISPR一词。
Cas9和PAM的发现
2005年5月,Bolotin正在研究嗜热链球菌的细菌,它刚刚被测序,揭示了一个不同寻常的CRISPR位点。尽管CRISPR阵列与先前报道的系统相似,但它缺少一些已知的cas基因,而含有新的cas基因,其中一个基因编码是他们预测具有核酸酶活性的大蛋白,即现在所知的Cas9。此外,他们还指出,与病毒基因有同源性的间隔区,在一端都有一个共同的序列。这个序列,原间隔基序(PAM)是目标识别所必需的。
自适应性免疫假设方案
2006年3月,Eugene Koonin通过计算分析研究一组同源蛋白质,并提出了一个假设方案,将CRISPR级联作为细菌免疫系统,其基于与噬菌体DNA同源的插入物,推翻了先前关于Cas蛋白可能包含一个新的DNA修复系统的假设。
自适应免疫的实验演示
2007年3月,科学家们想通过噬菌体来制造酸奶,并在乳制品工业中广泛使用。Horvath和他的同事通过实验证明了CRISPR系统确实是一个适应性免疫系统。此外,他们还发现Cas9可能是干扰所需的唯一蛋白质,CRISPR系统通过这个过程使入侵的噬菌体失活。
间隔子序列被转录为指导RNA
2008年8月,约翰·范德奥斯特等科学家们开始详细说明CRISPR-Cas系统是如何“干扰”入侵的噬菌体的。他们指出在大肠杆菌中,源于噬菌体的间隔序列被转录成小RNA,称为CRISPR RNAs,引导Cas蛋白质到达目标DNA。
CRISPR作用于DNA靶
2008年12月-Luciano Marrafini和Erik Sontheimer,他们证明了目标分子是DNA,而不是RNA。许多人认为CRISPR与真核生物的RNA沉默机制类似,后者以RNA为靶点。marrafini和Sontheimer在他们的论文中明确指出,如果这个系统能够被转移到非细菌系统中,那么这个系统将是一个强大的工具。
Cas9切割目标DNA
2010年12月,Moineau和他的同事证明CRISPR-Cas9在精确的位置,即PAM上游的3个核苷酸处,在靶DNA中产生双链断裂。他们还证实了Cas9是CRISPR-Cas9系统中进行切割所需的唯一蛋白质。
发现用于Cas9系统的tracrRNA
2011年3月,关于自然CRISPR-Cas9引导的干扰机制的最后一个难题来自Emmanuelle Charpentier。他们对化脓链球菌进行了小RNA测序,该链球菌具有含Cas9的CRISPR-Cas系统。他们发现除了crRNA外,还存在第二个小RNA,他们称之为反式激活CRISPR RNA(tracrRNA)。他们表明tracrRNA与crRNA形成双链体,而正是这种双链体将Cas9引导至其靶标。
CRISPR系统可以在其他物种中异源发挥作用
2011年7月,Siksnys和同事从嗜热链球菌(II型系统)克隆了整个CRISPR-Cas基因座,并在大肠杆菌(不包含II型系统)中表达了这一点,他们证明了它能够提供质粒抗性。这表明CRISPR系统是独立的单元,并证实II型系统的所有必需组件都是已知的。
Cas9介导的裂解的生化表征
2012年9月,利用异源系统, Siksny和他的团队从大肠杆菌携带嗜热杆菌CRISPR基因的大肠杆菌菌株中,用crRNA与crRNA复合纯化了Cas9,并进行了一系列生化实验,以表明Cas9的作用模式,他们验证了PAM的裂解位点和要求,并使用点突变,他们显示RuvC域切割非互补链,而HNH域切割互补位点。他们还指出,crRNA可以缩减到20 nt的伸展,足以有效切割。
CRISPR-Cas9用于基因组编辑
2013年1月,麻省理工学院麦戈文脑研究所的张峰在其他基因组编辑系统上工作,他是第一个成功地将CRISPR-Cas9用于真核细胞基因组编辑的人。张先生和他的团队设计了两种不同的Cas9同源序列(来自嗜热链球菌和化脓链球菌),并在人和小鼠细胞中展示了靶向基因组切割。从那时起,研究人员发现CRISPR/Cas9具有惊人的多功能性。科学家们不仅可以利用CRISPR将基因剪除,从而“沉默”,还可以利用修复酶将所需的基因替换到狙击手留下的“洞”中。

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 






 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 发表于 2022-8-30 10:02
发表于 2022-8-30 10:02