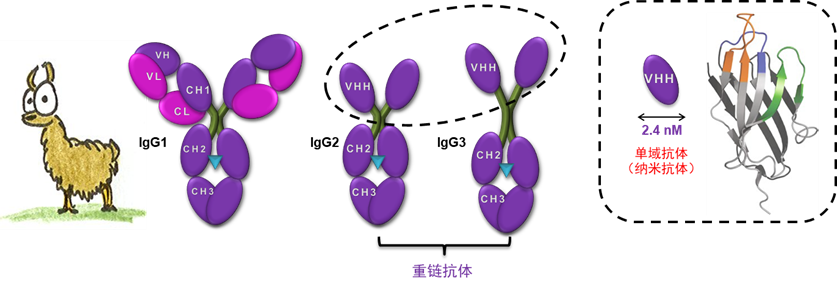
近年来,羊驼单域抗体(nanobody or VHH)的发展备受关注,成为生命科学界的热门研究领域。羊驼的单域抗体源自于羊驼独特的免疫系统,其小尺寸,高稳定性和出色的抗体抗原结合能力使其应用范围大为拓展,与传统抗体相比,单域抗体具有许多显著优势,使其在生物医学研究和临床应用中具有巨大的潜力。 背景:纳米抗体在GPCR研究中的应用 G蛋白偶联受体(G protein coupled receptor,GPCR),是细胞表面一类非常重要的受体蛋白,人类基因组中,GPCR编码基因所占比例超过2%,包含超过800个成员,是最大的一类细胞膜蛋白。据统计,目前已经有103种GPCR有临床使用的药物,针对多种疾病;目前正在开发的药物中,其中有142种药物是针对83种GPCR。目前已经有大量的G蛋白偶联受体有针对性的药物上市,并且有大量的药物处于正在开发的状态,GPCR是最有价值的药物开发靶点,未来会有更多的药物用来治疗各种疾病[1]。 靶向GPCR的小分子药物开发的同时,单克隆抗体药物的开发也受到了极大的关注,由于GPCR在胞外结构域比较小、信号激活比较复杂、传统IgG抗体由于本身结构比较大等原因,开发难度非常大,目前靶向GPCR的抗体药物开发并不顺利,已上市的抗体药物仅有2个( erenumab, 靶向CGRP-R 以及mogamulizumab,靶向CCR4),当前靶向GPCR的抗体药物急需以新的抗体形式开展[2]。 1993年,比利时科学家在自然杂志中首次报道羊驼外周血液中存在一种天然缺失轻链的抗体,它们的重链可变区(VHH区)负责抗原的识别与结合。更重要的是单独克隆并表达出来的VHH结构具有与原重链抗体相当的结构稳定性以及与抗原的结合活性,是目前已知的可结合目标抗原的最小单位,其分子量只有15KD,也因此被称作纳米抗体[3]。
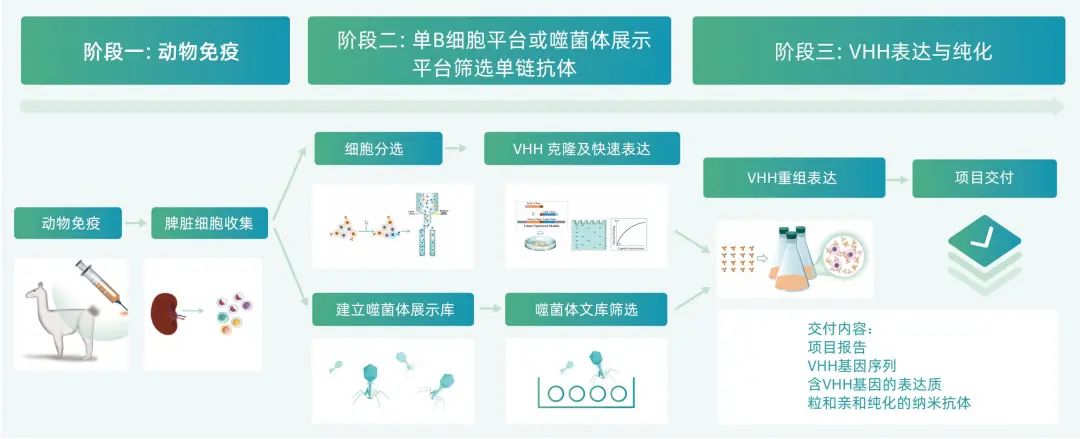
纳米抗体由于具有尺寸小,结构简单,肿瘤组织穿透能力更强和容易穿过血脑屏障,亲和力高,免疫原性低,亲水性好、稳定性高和易改造等特点,已广泛用于各项针对GPCR家族成员的研究。以纳米抗体的形式开发GPCR研究工具和抗体药物是未来的大趋势。当前针对GPCR,纳米抗体主要应用包括以下方面: GPCR的活性状态很不稳定,给结构解析的工作造成了很大的困扰。2011年,Brian K. Kobilka首次在nature在线发表nanobody识别β2AR激活构象并能稳定受体激活状态从而解析出β2AR在活性状态的结构[4]。这是一个里程碑式的工作,不仅解析出GPCR在活性状态下的结构,更提供了一种方法:nanobody可以在GPCR结构解析中发挥巨大的作用。在这之后,一系列的工作基于nanobody识别并稳定GPCR的活性状态在线发表,成功解析出一系列的GPCR活性状态的结构,包括US28[5](science,2015)、KOR[6](cell,2018)、AT1R[7](cell,2019)等,并在很大程度上推进了纳米抗体在GPCR研究领域的应用。 纳米抗体由于其高亲力和特异性,在GPCR细胞成像方面也得到广泛的应用,相比较与IgG抗体也具有明显的优势[8]。例如,在这篇文献中,科学家团队创造性地利用分别靶向,淀粉样沉积物,和细胞内Tau蛋白神经纤维缠结的特异性纳米抗体,在AD转基因小鼠模型中,通过静脉注射纳米抗体跨越血脑屏障,以及通过体内双光子成像显微镜,观测到AD转基因小鼠的病变过程,为神经学领域的诊断和治疗研究提供了新的工具。 Bryan L. Roth等2020在线发表文章中基于BRET技术,利用纳米抗体,可在细胞内实时检测受体的激活状态,主要是利用纳米抗体可以特异性识别GPCR的激活/非激活状态,将受体和纳米抗体分别带上不同的标签,根据在加药刺激前后BRET ratio的变化来实时检测受体的活性状态[9]。 GPCR内源性表达和状态检测 刘剑峰等2022年在Nature Chemical Biology报道了建立了基于纳米抗体的天然膜蛋白复合体表达与活性检测新方法,并发现代谢型谷氨酸受体(mGlu)二聚体在脑组织中高比例表达,包括mGlu同源二聚体和异源二聚体的分布和活性检测。纳米抗体由于其高特异性和高亲和力,在检测GPCR内源性定位、表达及聚合状态方面有独特的优势[10]。 目前已经有数款以纳米抗体为主要形式的治疗性药物被批准上市,包括Ciltacabtagene autoleucel,Calplacizumab和Ozoralizumab等。值得一提的是,在国产创新药公司当中,南京传奇生物的CAR-T疗法CARVYKTI通过纳米抗体实现靶向BCMA抗原,以及康宁杰瑞自主研发的全球首个皮下注射PD-L1药物也是一款以单域抗体为骨架的融合蛋白。 除此之外,在靶向GPCR的治疗性研究领域,有团队更进一步报道了能特异性识别GPCR受体激活状态的纳米抗体,2017年,Jean-Philippe Pin团队在nature communications发表靶向C族受体mGlu2筛选到一系列的纳米抗体,主要是识别mGlu2胞外VFT区,并且只识别受体激活状态等,筛选到的纳米抗体在能识别受体不同构象的同时,还具备激动剂效应和正向变构剂效应,这进一步说明纳米抗体不仅仅在GPCR结构解析中发挥着重要作用,在GPCR药物开发中也存在巨大的潜力[11]。之后,该研究团队也陆续报道了影响mGlu5活性的纳米抗体NB43[12];mGlu4的纳米抗体DN45[13];APJ受体的纳米抗体[14]JN241(受体抑制剂)、JN241-9(受体激动剂);AT1R受体的纳米抗体AT118i4[15] (受体抑制剂)等。 利用噬菌体展示平台开发GPCR纳米抗体 优睿赛思为国内首家基于单B细胞筛选技术和传统噬菌体展示技术的双平台羊驼单链抗体定制服务商。具备从抗原制备、动物免疫、抗体筛选到克隆,表达,表征和功能检测的完整服务链条,成功开发了一系列结合GPCR复杂结构膜蛋白的纳米抗体;同时,围绕纳米抗体公司具备多种检测方法,包括细胞成像、流式分析、HTRF等,确保成功交付满足终端应用需求。本团队建立的纳米抗体开发平台,主要优势在于: 独立知识产权单B细胞分选结合噬菌体展示技术,可提高项目成功率和纳米抗体的多样性 过表达GPCR靶点的细胞膜提取物作为抗原进行动物免疫 优化后的噬菌体展示载体,以及筛选方案 独特的算法 二代基因测序(NGS)结合独特的算法,从海量的数据中筛选出稀有的纳米抗体序列 美洲进口羊驼,本地养殖基地,保证无免疫背景的Naive羊驼动物 拥有大肠杆菌、毕赤酵母、哺乳动物细胞多种纳米抗体表达平台 基于以上平台优势,可以很高效筛选到高亲和力的纳米抗体,服务于GPCR的科学研究和纳米抗体药物的开发。 纳米抗体开发流程
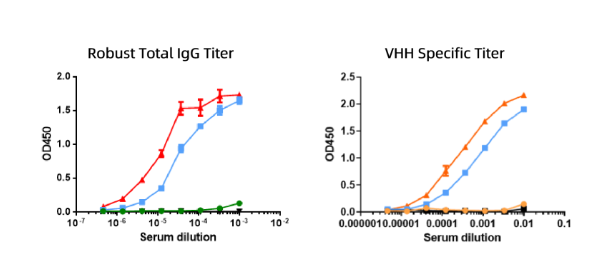
以某靶点为例,优睿赛思使用GPCR靶点稳定表达细胞株的细胞膜提取物进行羊驼免疫;在免疫过程中,利用该GPCR靶点过表达细胞和羊抗羊驼的IgG (H+L)二抗,通过细胞ELISA对免疫后血清进行总的IgG滴度以及VHH特异性滴度检测,从多个维度评估动物的免疫反应。(见图1)
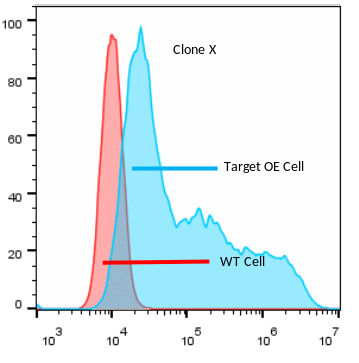
图1.血清效价检测 抽取的羊驼外周血建立的VHH文库构多样性为不低于109cfu,VHH插入率超过95%,经过培养后文库总容量约 1011cfu。以上述得到的纳米抗体文库为来源,利用该GPCR类靶点的细胞作为抗原,开展菌体表面展示筛选,用于后续流式分析实验研究。 从富集的产物中挑取表达有纳米抗体的微生物单克隆进行抗原特异性鉴定(流式细胞分析),最终获得35%阳性克隆可以和该GPCR类靶点的过表达细胞结合,具有流式功能(见图2)。同时,针对具有FACS克隆进行重组抗体可溶表达,有占比23%克隆可溶性表达较好。
图2.流式分析结果展示 最后,针对这这些克隆进行表位分析,发现有14%个抗体的表位在ECD1区,25%个抗体的表位在ECD2区,36%个抗体的表位在ECD3区,还有25%个抗体的表位未知。说明开发的抗体的多样性较好。(见图3)
图3.阳性克隆结合表位分析 综上所述,利用该噬菌体展示平台进行羊驼纳米抗体开发成功获得了高特异性、高亲和力和多样性较好的纳米抗体,具体流程为:
基于独立知识产权单B细胞分选技术,并结合深度优化的噬菌体展示技术,以及在纳米抗体发现领域深耕累积出来的优势,优睿赛思热切期待与整个行业的仁人志士开展深度合作,通过共同努力,进一步挖掘纳米抗体的潜能,加速纳米抗体的技术发展,推动生命科学的进步,并为全世界人类的健康事业做出贡献。 有意请填写表单: https://jinshuju.net/f/LIixhg 或点击文章左下角“阅读原文”填写 参考文献 [1] Congreve M, de Graaf C, Swain N A, et al. Impact of GPCR Structures on Drug Discovery[J]. Cell, 2020,181(1):81-91. [2] Catherine J. Hutchings. Mini-review: antibody therapeutics targeting G protein-coupled receptors and ion channels. Antibody Therapeutics, 2020, Vol. 3, No. 4 257–264 [3] Hamers-Casterman, C., Atarhouch, T., Muyldermans, S., Robinson, G., Hamers, C., Songa, E.B., Bendahman, N. and Hamers, R. (1993) Naturally occurring antibodies devoid of light chains. Nature, 363, 446-448. [4] Rasmussen S G, Choi H J, Fung J J, et al. Structure of a nanobody-stabilized active state of the beta(2) adrenoceptor[J]. Nature, 2011,469(7329):175-180. [5] Burg J S, Ingram J R, Venkatakrishnan A J, et al. Structural biology. Structural basis for chemokine recognition and activation of a viral G protein-coupled receptor[J]. Science, 2015,347(6226):1113-1117. [6] Che T, Majumdar S, Zaidi S A, et al. Structure of the Nanobody-Stabilized Active State of the Kappa Opioid Receptor[J]. Cell, 2018,172(1-2):55-67. [7] Wingler L M, McMahon C, Staus D P, et al. Distinctive Activation Mechanism for Angiotensin Receptor Revealed by a Synthetic Nanobody[J]. Cell, 2019,176(3):479-490. [8] Li T, Vandesquille M, Koukouli F, et al. Camelid single-domain antibodies: A versatile tool for in vivo imaging of extracellular and intracellular brain targets[J]. Journal of Controlled Release, 2016,243:1-10. [9] Che T, English J, Krumm B E, et al. Nanobody-enabled monitoring of kappa opioid receptor states[J]. Nature Communications, 2020,11(1). [10] Meng J, Xu C, Lafon P A, et al. Nanobody-based sensors reveal a high proportion of mGlu heterodimers in the brain[J]. Nat Chem Biol, 2022. [11] Scholler P, Nevoltris D, de Bundel D, et al. Allosteric nanobodies uncover a role of hippocampal mGlu2 receptor homodimers in contextual fear consolidation[J]. Nat Commun, 2017,8(1):1967. [12] Koehl A, Hu H, Feng D, et al. Structural insights into the activation of metabotropic glutamate receptors[J]. Nature, 2019,566(7742):79-84. [13] Haubrich J, Font J, Quast R B, et al. A nanobody activating metabotropic glutamate receptor 4 discriminates between homo- and heterodimers[J]. Proc Natl Acad Sci U S A, 2021,118(33). [14] Ma Y, Ding Y, Song X, et al. Structure-guided discovery of a single-domain antibody agonist against human apelin receptor[J]. Sci Adv, 2020,6(3):x7379. [15] McMahon C, Staus D P, Wingler L M, et al. Synthetic nanobodies as angiotensin receptor blockers[J]. Proc Natl Acad Sci U S A, 2020. Yurogen 公司简介 Yurogen Biosystems (优睿赛思)2015年成立于美国波士顿,并于2018年在武汉光谷设立优睿赛思(武汉)生物科技,服务于亚太市场。2021年优睿赛思生物科技和爱博泰克生物科技(ABclonal Inc.)实现了合并,并成为ABclonal的全资子公司。 |

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号