主要而言,引物的大小约为20个核苷酸,与靶点完全互补,经常用于PCR。然而,确实存在很多为特殊应用设计的引物变异。例如,这些引物变异可以更长,可以拥有特殊的功能、错配、标签或碱基替代物(图1)。它们可以为识别特殊的突变(如易位)而设计,也可以满足特殊的标准(如在多重配置中没有交叉杂交)。
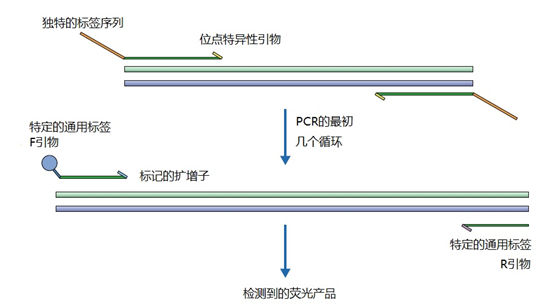
图1 | 不同类型的引物 a 最小引物:引物,与靶点不完全匹配(也叫变性引物)。错配并不总是像预期的那样影响杂交体的稳定性。在某些错配中存在对稳定性的一些贡献。 b 外加引物:一个带有5′外挂的引物,以便将额外的序列附加到扩增物上。该序列不与本地靶点退火。这个延伸序列通常用一个间隔物与5′末端的序列相连。这个延伸序列也将在随后的循环中被扩增(见图2)。一个额外的功能被添加到扩增物中,如用于克隆的限制性位点或用于蛋白质合成的ATG起始密码子,扩增物作为一个基质发挥作用。 1、扩展引物 扩展引物在5'端含有额外的核苷酸,不与靶点杂交,在第一个PCR循环中不被复制。这些引物在第二个PCR循环后成为扩增物的一个组成部分。通过这种方式,可以添加额外的序列,如限制性位点、翻译起始序列或启动子序列,最多为50个碱基。 一种特殊的扩展引物类型配备了一个编码标签,其序列与问题相适应。这个标签可以作为标签互补引物或探针的杂交靶点(图2)。
图2 | 带有5′-外挂的标签杂交原理 在基因座特异性引物对上添加一个5′标签。该标签由基因组上不存在的序列组成。当标签成为扩增物的一部分时,就形成了一个新的靶点序列。一个带标签的特异性引物将接管扩增反应。标签有助于在与正向引物耦合的荧光色素的基础上进行识别。MLPA(见图4)使用标签扩增的原理。 标签也可以被设计成显示出独特的能力,例如形成发夹环。发夹引物利用这种构造来猝灭非退火引物的信号。扩增后,标签将最终成为扩增物中的一个线性序列。这样一来,猝灭剂和荧光剂就会分离,从而导至荧光的产生(图3a)。同样的原理适用于猝灭的探针(图3b)。有些标签有独特的序列,在自然界中不存在。通过使用多个标签,可以同时捕获多个扩增物。
图3 | 实时PCR中引物和探针的猝灭 引物(a)和探针,即分子信标(b)在溶液中不发亮,这是猝灭的结果。报告基团和猝灭者在彼此附近,从而交换激发电子(猝灭)。一旦发夹引物在PCR过程中被拉长(a)或探针与扩增物退火(b),荧光剂和报告基团将被物理分离并发生荧光。 多重连接依赖性探针扩增(MLPA)可使用另一种类型的扩展引物(图4)。
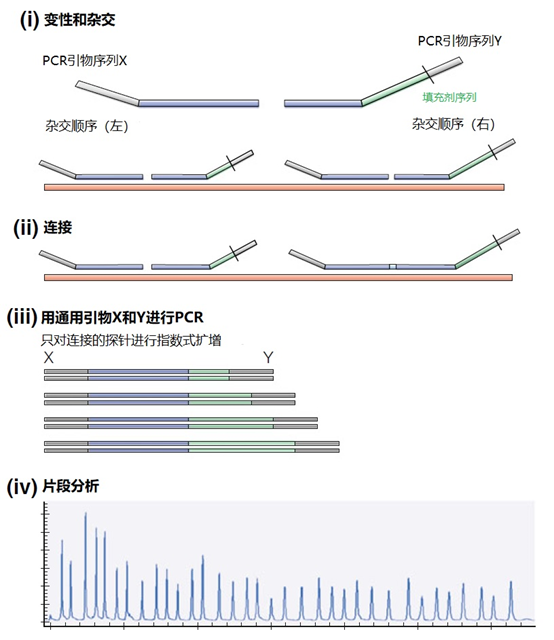
图4 | 多重连接依赖性探针扩增(MLPA) 该技术中使用了标签(见图2)。反应从双重杂交开始(探针的蓝色部分)。两个探针以头对尾的构象同时与靶点杂交。连接酶连接两个探针的 3'-5'-末端。标签序列 X 和 Y 以及填充序列(S)不杂交。每个探针组具有不同的唯一填充序列长度(S1、S2、S3 等)。这种独特的长度便于识别每个放大器。所有连接的探针都用靶向序列 X 和 Y 的相同引物对进行扩增。通过毛细管电泳区分扩增子的填充长度。 MLPA是一种PCR技术,两个探针在靶点上头尾相接地杂交。独特的标签位于两个探针的3ʹ(右)或5ʹ(左)侧。两个标签都包含一个通用引物序列(X和Y)。此外,在5'侧有一个特殊的标签,即所谓的填充序列。 这个缓冲序列的长度是可变的。引物和填充物的序列没有相互作用,如果需要,可以使用几十个不同长度的序列。在靶点杂交后,相邻的探针被连接起来。现在,可以使用与X和Y互补的引物与悬吊的标签退火,进行PCR。合成的扩增物(对10-30个不同靶点的特异性)可以通过(毛细管)电泳和/或浓缩来分离(见图5和图6c)。
图5 | 多重连接依赖探针扩增法(MLPA) a 用于检测靶点序列A和B的MLPA法原理。两个完全匹配的(半)探针与靶点A或靶点B杂交,每个探针都有一个带有3′引物退火位点的标签(上游)。下游不仅有一个引物退火位点,而且还有一个所谓的填充序列,其长度可以变化。杂交后,探针通过一个5′-3′连接酶的磷酸糖桥进行共价连接。这样就产生了一个新的靶点序列,由引物退火位点、独特序列和填充序列组成。在所有连接的探针变性后,开始进行PCR。由于两个半探针的连接只发生在3′-5′的完美匹配处,PCR产物也只有在这时产生,这决定了这种方法的高选择性。不同填料的长度差异使得产品可以按大小进行分离。 b 毛细管电泳对PCR产物的分辨率足以检测同源性或杂合性,或者可以检测到完全缺失。第二行是BRCA1基因外显子13缺失的患者与正常质控组的对比。
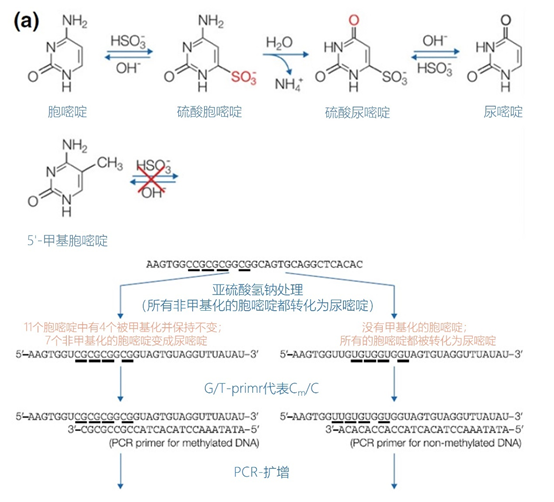
图6 | 甲基化特异性PCR:原理和三种不同的检测方法 三种方法中的每一种都需要对胞嘧啶进行亚硫酸氢盐处理并将C转化为U。这样可以区分甲基化的CpG位点(保留C)和非甲基化的胞嘧啶(变为U)。 b MS-MLPA(甲基化特异性MLPA),可以检查40个位置的甲基化状态。在亚硫酸氢盐处理后的两个反应中使用两个探针。左边是一个带G的半探针的例子,可以检测到C没有变化的甲基化靶点,右边是一个带A的半探针的例子,可以识别非甲基化的位置。就像图5中一样,填料序列的长度不同。在毛细管电泳后,可以确定(副产品的长度)是否涉及同型、异型或野生型样品。 c 传统的PCR,有两种不同的测定。在「C」在亚硫酸氢盐处理后未被甲基化的位置,「C」在亚硫酸氢盐处理后未被甲基化。 在亚硫酸氢盐处理后「C」未被甲基化的位置,「C」已被转化为「U」。引物,一种是带有G/A取代的碱基的测定,另一种是带有原始「G」的引物,将选择性地结合。如果有甲基化的部分,用含有G的引物进行PCR。当C被转化为U时,用另一个引物(用A代替G)进行PCR。PCR本身是用普通的dNTPs完成的。 也可以应用一个能与新引物杂交的序列;如扩增片段长度多态性(AFLP)技术(见图7)。
图7 | 扩增片段长度多态性(AFLP) AFLP是通过扩增不同的限制性片段来获得全基因组的图像,随后通过毛细管电泳进行分析。片段的模式可以是物种、类型或亚类型特定的 首先,用限制性酶处理基因组,造成两边都有粘性末端的双链片段。使用常用切割器(作用于相对常见的序列,如EcoRI)和罕见切割器(相对罕见的序列,如MseI,TaqI)的组合。这两种酶之间的比例决定了产生的片段数量 (a) 互补适应体与「粘性末端」杂交。使用连接酶(b)将适配体和限制性位点连接到基因组DNA上。通过这种方式,DNA被扩展成可以结合互补引物的通用序列(c)。此外,引物可以用1-3个额外的碱基进行扩展,限制了PCR产物的数量。在c中,引物被提供了三个额外的碱基(在此图中「GTA」代表MseI-引物,「AGC」代表EcoRI-引物)。完全(100%)的互补性将只出现在3′侧有限的地方;只有这样,引物才会被扩展为扩增物。
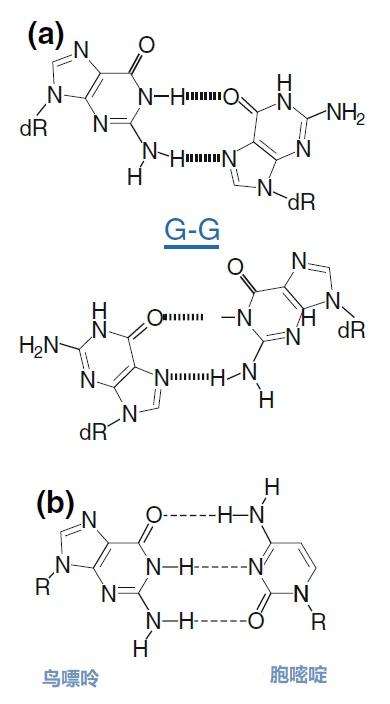
图8 | 37种HPV类型和3种质控的简化核苷酸序列比对 从病毒性HPV基因组的6582至6646位进行HPV类型的核苷酸序列比对 正向(SPF1)和反向(SPF2)用绿色矩形表示。37种HPV类型之间发现不同碱基的位置用「N」表示,排列中的同源性用「-」表示。大多数序列有3-5个不明确的地方。正向和反向序列位置的3′侧具有大多数同源性。针对HPV16的引物SPF1/2将导至完全匹配。引物中6592和6595位置的肌苷的吸收避免了与其他类型的不匹配。引物浓度和PCR条件的选择是为了使引物能与其他类型的HPV进行交叉杂交。 位置6601和6624之间的区域(黄色矩形)是高度可变的。它包含类型特异性探针的热点,可以区分使用SPF1/2引物对合成的所有37种HPV类型的扩增子。(关于碱基组成,见图2) 由于3'端的核苷酸差异阻碍了PCR的进行,引物的设计需要将5'端的非互补段与3'端的完全匹配放在一起。也可以设计一个引物,在有碱基差异的位置上有一个随机碱基或SNP的特殊组合特征。例如,「R」(A或G),「B」(C或G或T)或「N」(任何碱基);正确的字母可在IUPAC/FASTA代码中找到。 退化的引物通常较长,以补偿杂交能力的损失。显然,错配不是绝对的;G-G、T-T、A-A和C-C之间的任何H键都有可能。然而,这将导至螺旋结构的变形和不太稳定的杂交体(图9)。
特别是在微生物研究中,以及在淋巴瘤/白血病的诊断中,可能需要用一对引物检测多个靶点(例如,使用16S rRNA基因上的类似序列的若干变异基因型的病毒或多个细菌物种)。这种引物被称为共识引物;但实际上,这些引物也被称为退化引物。 可以用肌苷替换引物中某些关键位置上的错配核苷酸。这个碱基自然存在于tRNA中,并与A、U和C化学上「匹配」。因此,肌苷可以与某些错配杂交,但对特异性没有任何贡献。 3、转位引物 转位引物的设计是为了在转位部位的上下游进行杂交。最为人所知的是恶性肿瘤的易位,其中有些肿瘤具有特定的亚型,适合于分子诊断。 在易位时,不同的染色体重新组合成一个完整的新序列。易位引物会在断裂点的两边进行杂交,从而确保只发生与易位有关的扩增。它们不会扩增没有易位的野生型序列。 4、多重PCR 在多重PCR策略中,可以在一个PCR反应中同时检测多个靶点。多重PCR使用2对以上的引物,在一个PCR反应中实施5对甚至10对引物,可以有效(或适当)使用试剂和仪器;相对降低了成本,减少了工作量。用于多重PCR的引物符合对普通引物有效的相同规格。 此外,所有可能的PCR反应将在相同的条件下进行,相当于反应间的差异。此外,与其他每个引物形成双链的风险必须是最小的。要求与靶点有同样强的结合力(相同的Tm),也要求每个单独的单倍体PCR反应有相当的效率。由于某些引物在与其他引物形成双倍体后可能出现假阴性,导至无法与靶点退火,因此避免这种双倍体的形成至关重要。关于这最后一个标准,引物通常要长一些,需要非常仔细地选择所需的引物浓度(见图10)。
图10 | 优化多重设计的阶段和可能性。传统的PCR需要PCR后的步骤,如电泳和杂交。实时PCR方案的开发始于使用插层染料,如SYBR® Green,并结合熔解曲线分析。反应效率将使用参考曲线确定。下一阶段是对探针的优化。 多重方法在微生物学中经常被用来从活检、拭子、尿液、粪便或其他样品中的许多可能的病原体中挑选出一个。另一个应用是血液学恶性肿瘤的基因定量。 对于所有的多重PCR方案,强烈建议首先在干试验中测试所有的引物,以估计引物在单倍体PCR反应中的表现以及随后在多重PCR格式中的表现。此外,还必须在实践中使用适当的质控样品测试所有引物在多重PCR方案中的正确活性。 5、重复引物 在大多数诊断PCR反应中,一个引物对用于扩增一个独特的靶点序列。少数诊断引物被设计成在基因组上的多个位置与相同的靶点杂交,并在同一试管中为一些不同的PCR反应提供引物。穿插重复序列,如广泛存在的简单序列重复(SSR)包含长段的简单序列。 一个著名的小型SSR的例子是(CACA)n-repeat和它的引物与重复的(GTGT)m。由于存在许多重复,引物在所有这些位点退火,初始化DNA合成将在大约40个PCR循环后产生不同大小的扩增物池(见图11)。
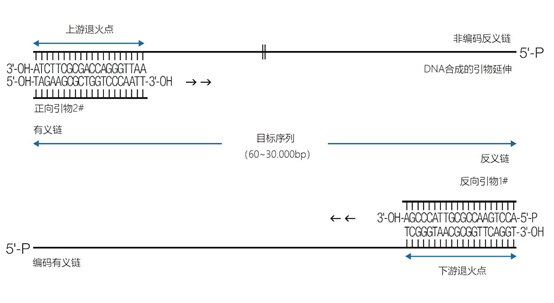
图11 | PCR中的引物退火点、合成方向和靶点序列 该图中描述了最重要的术语。传统PCR的靶点通常为60-400个碱基;而实时PCR的靶点为80-150个碱基。 这种「重复间」PCR中扩增物的电泳模式,即所谓的指纹,是基因组特异性的,可用于基因分型。 同样的原则也可适用于大中型重复序列(10-300 bp),如ERIC重复序列和各种BOX序列,这些序列经常用于微生物学分型。 |

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号