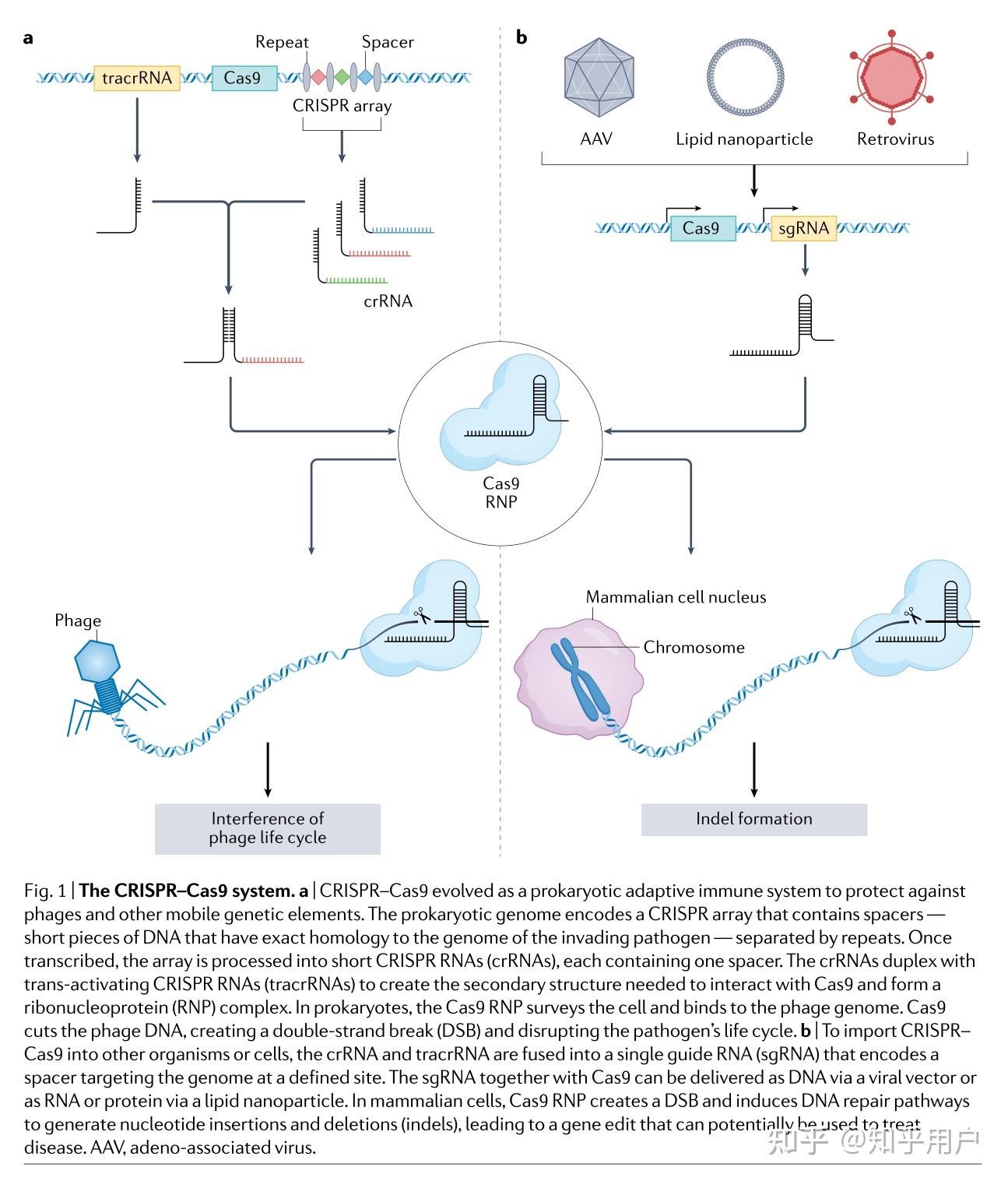
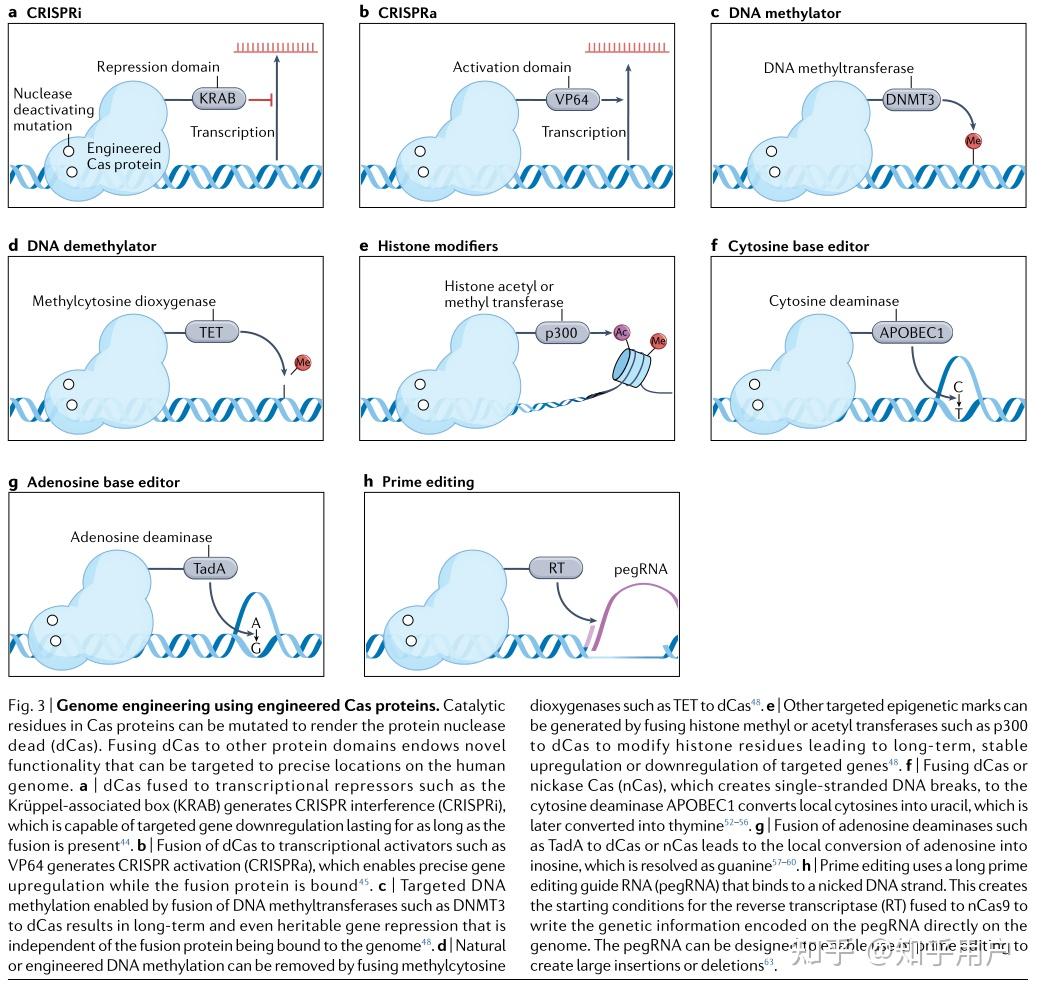
CRISPRa和CRISPRi基因调控方法可以实现短期的基因调节。在有丝分裂后的细胞或需要短暂基因表达调控的疾病中,这种短暂性并不构成问题。然而,一些疾病需要长期且可遗传的基因调控变化。通过向DNA添加甲基基团或在组蛋白残基上插入乙酰基或甲基基团,可以在局部调控基因表达的同时进行表观遗传修饰。这些修饰通常具有持久性,并且可以通过子细胞传递,为长期的基因表达调控提供了机会。许多表观遗传修饰因子已经与CRISPR蛋白融合,以在DNA或染色质水平上实现化学修饰。例如,CRISPRoff和CRISPR-KAL通过修饰H3K9me3和DNA甲基化实现了长期(数月)的基因沉默。这些方法可能适用于需要持续基因调控的疾病治疗。
将来自DNMT3家族的DNA甲基化结构域与dCas9融合,可以实现长期的基因沉默。例如,将dCas9-DNMT3融合蛋白靶向SNCA内含子1,在携带SNCA三倍体的人类iPSC来源的多巴胺能神经元中生成靶向DNA高甲基化,从而在体外挽救了帕金森病相关的表型。为了逆转自然DNA甲基化的沉默效应,将十一转位甲基胞嘧啶二氧化酶1(TET1)催化结构域与dCas9融合,可以选择性地去除DNA甲基基团并上调基因表达。这种方法已被研究作为治疗脆性X综合症的潜在疗法,该综合症是由FMR1基因中CGG重复扩增引起的智力障碍,导致广泛的甲基化,从而降低基因表达。将dCas9-TET1靶向FMR1可以去甲基化CGG重复序列,重新激活持久的基因表达,并在体内iPSC来源的神经元中挽救疾病表型。包含dCas9和TET酶催化结构域的融合蛋白也已被用于体外治疗癌症(靶向BRCA1)和体内治疗(靶向SARI),并在体内减轻肾脏纤维化(靶向Rasal1或Klotho)。重要的是,由此产生的DNA甲基化变化是持久的、可遗传的和可逆的。
为了定点修饰组蛋白,p300结构域的催化核心被融合到dCas9上。当指向DNA上的增强区时,这种融合会在组蛋白H3的赖氨酸27上增加一个乙酰基团(H3K27ac),导致基因表达的激活。在小鼠中,dCas9-p300的表达能够上调T细胞中Foxp3的表达,将其转化为调节性T(Treg)细胞,具有治疗自身免疫的潜力。使用与组蛋白去乙酰化酶1(HDAC1)融合的dCas9可以去除H3K27ac标记。这种方法已经针对KRAS来抑制癌症生长。其他抑制性CRISPR组蛋白修饰剂包括减少H3K4甲基化,增加H3K9甲基化和增强HP1α结合,当靶向GRN时,可以减少肝癌细胞的增殖和侵袭。
Base and prime editing
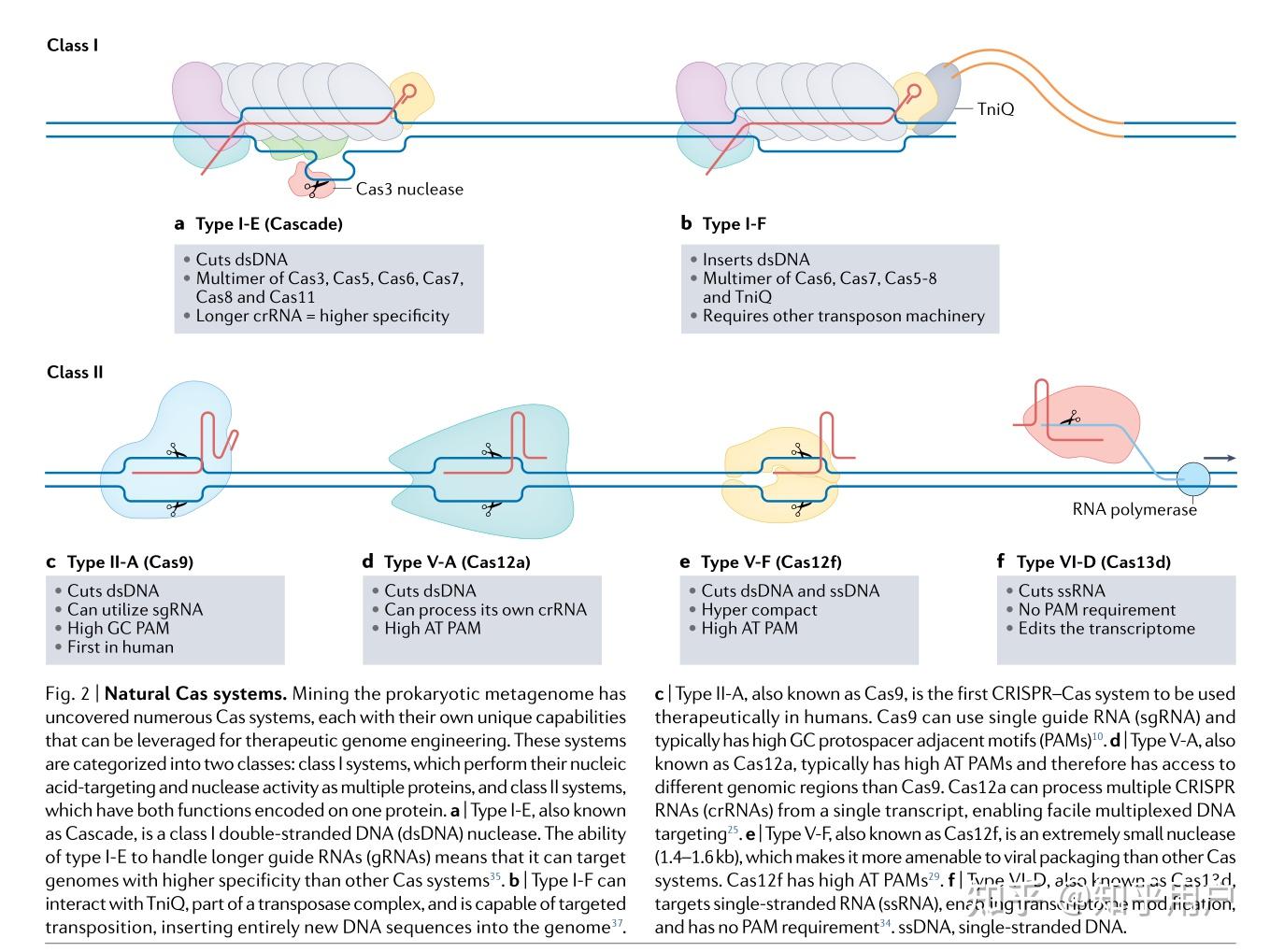
除了细胞疗法外,野生型CRISPR系统往往是精确突变纠正的不良选择,因为添加的核苷酸的数量或特性无法控制。为了填补这一空白,已经产生了能进行精确基因改变的CRISPR融合形式,并在无数种疾病中得到应用。
Base editing
随着技术的迅速进步和对有害点突变进行无与伦比精确修复的能力,碱基编辑技术(base editors)被迅速应用作为治疗已知错义突变相关的明确疾病的潜在方法。能够创建C到T突变的CBE已被广泛用于各种体内模型中。Cas9和Cas12a的CBE已被用于修复Pah基因的错义突变,该基因存在于人类常染色体隐性肝脏疾病苯丙酮尿症(PKU)的小鼠模型中。C到T转换的能力使得产生终止密码子成为可能,终止密码子总是以胸腺嘧啶(thymine)开头。在肌萎缩侧索硬化症(ALS)的小鼠模型中,使用SpCas9 CBE在SOD1基因中创建一个过早终止密码子,减少了肌肉萎缩并改善了神经肌肉功能。由于CBE CRISPR构建的体积较大,需要将蛋白质分割成两个AAV载体,并在细胞内使用肽内切酶进行翻译后融合。
ABEs与治疗方法高度相关,因为在所有已知的致病点突变中,C-G到T-A的转换约占一半。在杜氏肌肉萎缩症小鼠模型中,以两个AAVs形式传递到肌肉的ABEs能够纠正杜氏肌肉萎缩症的一个突变,并改善疾病表型。ABEs也被用来纠正哈钦森-吉尔福德早衰综合症小鼠模型中的LMNA突变,将中位寿命从215天延长到510天。值得注意的是,使用ABE来纠正镰状细胞病小鼠模型中的无义突变,导致该疗法的BEACON-101 I/II期试验获得批准。基因编辑的其他治疗用途已在其他地方进行了回顾。
Prime editing
Prime editing具有广泛的治疗基因组编辑潜力,但与其他CRISPR系统相比,其研究尚不如其他方法广泛。在首次描述该技术的研究中,研究人员成功纠正了导致镰状细胞贫血症的HBB基因突变、导致Tay-Sachs病的HEXA基因突变以及保护免疫性蛋白疾病的PRNP基因。Prime editing能够进行精确的基因突变,这是使用基因碱基编辑器目前无法实现的。例如,在α1-抗胰蛋白酶缺乏症(AATD)小鼠模型中,研究人员通过创建A到G的基因编辑,成功将prime editing技术应用于小鼠肝脏中,纠正了SERPINA1基因中的致病性E342K突变。此外,研究人员还利用prime editing技术在小鼠视网膜中纠正了Dnmt1基因的突变,通过创建G到T的基因转换,展示了该技术纠正眼部疾病的潜力。这些精确的基因编辑通过其他CRISPR工具无法实现。
除了基因碱基编辑,prime editing还可以用于插入寡核苷酸。在人类诱导多能干细胞(iPSCs)中,利用prime editing技术将两个核苷酸(AC)插入到DMD基因的第52外显子中。这种方法使得外显子重构可以恢复DMD的表达,并改善模拟杜氏肌营养不良症的iPSC-来源心肌细胞的收缩功能。尽管prime editing技术在治疗上仍处于起步阶段,但这种方法创造的基因组编辑的灵活性潜在地能够纠正许多疾病。
感染的预防和治疗

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 






 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 发表于 2025-5-25 12:05
发表于 2025-5-25 12:05