用户名
UID
Email
密码
记住
立即注册
找回密码
只需一步,快速开始
微信扫一扫,快速登录
开启辅助访问
快捷导航
门户
Portal
社区
BBS
资讯
会议
市场
产品
问答
数据
专题
帮助
签到
每日签到
企业联盟
人才基地
独立实验室
产业园区
投资机构
检验科
招标动态
供给发布
同行交流
悬赏任务
共享资源
VIP资源
百科词条
互动话题
导读
动态
广播
淘贴
法规政策
市场营销
创业投资
会议信息
企业新闻
新品介绍
体系交流
注册交流
临床交流
同行交流
技术杂谈
检验杂谈
今日桔说
共享资源
VIP专区
企业联盟
投资机构
产业园区
业务合作
投稿通道
升级会员
联系我们
搜索
搜索
本版
文章
帖子
用户
小桔灯网
»
社区
›
C、IVD技术区
›
基因编辑技术
›
基因编辑与载体
图文播报
2026庆【网站十三周
2025庆【网站十二周
2024庆中秋、迎国庆
2024庆【网站十一周
2023庆【网站十周年
2022庆【网站九周年
返回列表
查看:
5858
|
回复:
0
[分享]
基因编辑与载体
[复制链接]
检验之星
检验之星
当前离线
金桔
金币
威望
贡献
回帖
0
精华
在线时间
小时
发表于 2025-4-24 19:57
|
显示全部楼层
|
阅读模式
登陆有奖并可浏览互动!
您需要
登录
才可以下载或查看,没有账号?
立即注册
×
基因编辑是老生常谈的内容,包括基因敲除(knockout,KO),基因敲低(knockdown,KD)和基因敲入(knockin)。在考研的时候,只是粗略涉及到了相关概念,具体的流程和注意事项并没有细看,今天看论文涉及到了基因编辑内容以及AAV(腺相关病毒)作为载体的基因编辑内容,因此在此总结学习一下。
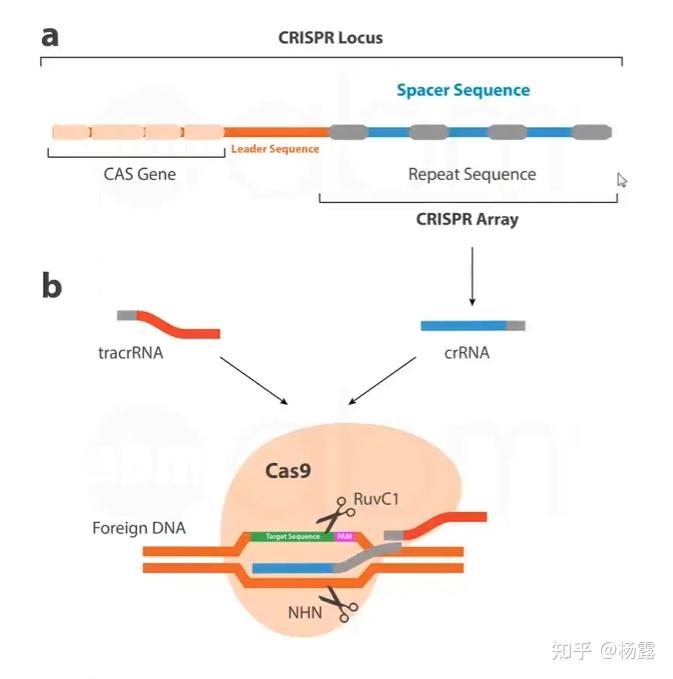
基于CRISPR/Cas9的基因编辑系统
CRISPR/Cas9系统最早在细菌和古细菌中发现的,属于一种免疫应对方式。外来DNA被核酸酶切割,然后以间隔序列的形式被被保守的重复序列捕获并整合到crispr位点,所得到的间隔片段作为创建crRNAs(创建短CRISPR RNAs,creat short CRISP RNAs)的模版,crRNAs与tracrRNA(反式激活cRNA,trans-activing crRNA)形成复合物,起到引导链的作用,将Cas9酶引导至互补的入侵DNA进行结合切割。
细菌体内Cas9酶识别切割外源DNA
在细菌识别外源DNA的时候,
PAM序列
起到了很大的作用。PAM序列是靶基因下游的一段短的脱氧核糖核苷酸序列(上图粉色部分,通常2-6个碱基),在gRNA或者复合物与Cas9酶结合后,Cas9酶会扫描外源DNA是否有对应的PAM序列,如果存在,解旋DNA,gRNA或者复合物识别并结合对应的DNA序列形成RNA-DNA结构,Cas9酶开始工作(通常在PAM序列上游切割);否则不会产生切割。PAM序列的存在保证了细菌自身的位点不会被切割。
那么,为什么靶位点的下游通常有PAM序列以及细菌第一次是怎样识别切割外源DNA的呢?实际上,在第一次切割外源DNA形成间隔片段的时候,是Cas1-Cas2参与切割捕获的。以噬菌体为例,当噬菌体入侵细菌时,其DNA会暴露在细菌胞质中,并被细菌的非特异性DNA监测系统(如RecBCD复合物)降解成片段,Cas1-Cas2复合物会从外源DNA中截取一段序列(称为原间隔序列,protospacer),这个序列必须满足两个条件:原间隔序列旁侧必须有PAM(如SpCas9对应的NGG);截取的protospacer长度通常为20-30bp(与后续gRNA长度匹配)。Cas1-Cas2会将protospacer(含PAM)插入细菌CRISPR阵列,但只保留protospacer本身,PAM不会被整合。因此实现了细菌第一次识别切割外源DNA并且以后的Cas9酶可以更轻松的识别切割对应的外源DNA。
gRNA
指的是工程化的单链嵌合体RNA,其综合了tracrRNA的支架功能和crRNA的特异性。不同菌株和不同类型的CRISP-Cas9系统对应的PAM序列不同,对于常用的化脓链球菌Cas9(
SpCas9
),PAM位于sgRNA靶向序列的3'端(下游),序列为NGG(N为任意碱基,G为鸟嘌呤)。
在Cas9酶切断了靶基因的DNA双链后,生物体肯定不会只是看着自己的DNA被外来的一把剪刀剪断后就不管了,肯定会进行DNA的断裂修复。在这里,DNA双链断裂后的修复分为了非同源末端连接修复(NHEJ)和同源定向修复(HDR)。基因敲除用到了前者,而基因敲入则用到了后者。在DNA被Cas9切断双链后,可以进行
NHEJ修复
,将两个断裂末端连接,而在连接过程中碱基在聚合酶和核酸酶作用下随机去除与添加,最终修复的基因组与原基因组相比有少量碱基的插入与缺失,类似于产生了移码突变,从而导致基因无法表达,即敲除(其实这里也有不少学问,比如在设计gRNA时,考虑到这个类似于移码突变的原理,最好将位点选在启动子或者编码抗体结合位点或者前几个外显子的基因片段上,这样移码突变万一产生3的倍数个碱基的变化也能实现敲除效果,同时也避免了WB可能的假阳性结果);同时,也可以进行
HDR修复
,随着Crispr/Cas9递送的还有修复模版(Donor DNA),其中间是要敲入的目的基因,两臂与被切位点两端有着长的相同的序列(同源臂),从而可以充当修复模版,实现基因定点敲入(其实这里有更多学问,比如如何提高敲入成功率,需要抑制NHEJ修复,或者让gRNA与修复模版结合在一起)。
在实际运用中,只要掌握了原理,方法还是很多的。比如,我可以将RNP复合物通过电转方式递送到细胞内实现基因编辑;可以运用质粒载体,运载Cas9基因和gRNA进行转染;进一步的,我可以用双荧光质粒载体,一个用来运送Cas9和gRNA以及红色荧光,一个运送修复模版和绿色荧光,用流式细胞仪筛选敲入的细胞;甚至我自己都能想一个出来,构建带有肝脏特异性启动子的Cas9基因小鼠,然后再注射肝脏靶向的有gRNA的AAV病毒载体,双重保险,实现肝脏部位的基因特异性敲除(这个我感觉很像Cre/Loxp系统,虽然我也不知道有啥区别,但我这个不也挺高效的,感觉唯一难点是不是构建带有肝脏特异性启动子的Cas9基因小鼠,同时脱靶风险高?)。
除了经典的基因敲除和敲入,还有dCrispr系统。顾名思义,dCrispr系统指的是死掉的(失活的)Crispr酶。dCrispr没有了DNA酶的活性,但仍有特异性识别和结合DNA的能力,再与其他一些抑制蛋白结合使用,从而在DNA水平上实现了基因表达抑制,使得高度特异性抑制非编码RNA,miRNA和核定位RNA成为可能;相应的,与其他一些激活蛋白结合使用,可以激活基因表达。
张峰老师作为这把基因剪刀的发明人,设计了一大坨方法来进行基因编辑。但万变不离其宗,基本原理还是相同的,唯一值得注意的是Crispr/Cas9实现基因敲入的过程,涉及到Donor DNA。
在此列举几个比较详细的实操文章,以供参考:质粒载体的
基因敲除
与
基因敲入
;
基于Cre/Loxp的基因编辑系统
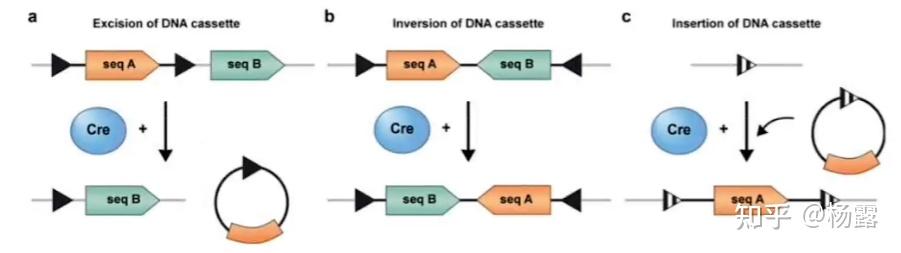
Cre/Loxp系统包括Cre重组酶和Loxp位点。Cre重组酶最早在P1噬菌体中被发现,其可以特异性识别Loxp位点,介导两个Loxp位点之间的特异性重组,使得Loxp位点间的基因被重组或修饰。Loxp(Locus of crossing over (x) of P1)位点最早也是发现于该噬菌体中,由两个13bp的反向重复序列和8bp的间隔序列组成,间隔序列决定的Loxp位点的方向。
经典Loxp位点
Loxp位点的位置和方向决定了Cre重组酶的作用方式,也决定了位点间基因的最终去向。
Loxp位点的位置和方向决定位点间基因的最终去向
由上图可知,Cre/Loxp系统可以用于基因敲除,基因敲入等编辑操作。在实际操作中,通常是先构建含有Loxp位点的小鼠(你想操作的基因,杂合纯合均可,称为Flox小鼠)和带有Cre酶基因的小鼠(可以在Cre酶基因前加上组织特异性启动子等操作实现空间特异性表达),然后杂交,筛选带有Cre酶基因的含有纯合Loxp位点的小鼠,从而实现了基因编辑。除此之外,还可以将Cre酶和比如性激素受体结合,注射外源性激素时激活受体Cre酶可以发挥作用,从而实现时间上的特异性表达(大意如此)。
多么复杂!光是杂交和筛选我估计都得大半年时间,实际上这也是Cre/Loxp系统的不便之处,编辑周期长,甚是不便,所以个人感觉,在实际运用中,除了那种敲了致死的基因,基本上还是Crispr/Cas9用得多。
基于双机制的基因编辑编辑系统
既然已经有了高效的基因剪刀Crispr/Cas9系统,为什么还需要Cre/Loxp系统呢?这就不得不提到两者的区别和联系了。
Cre/Loxp系统可以实现时空特异性的条件基因操作:Cre/Loxp系统可以实现在模式动物的某个特定的发育阶段或者某个特定的组织或器官进行基因敲除,避免了全身基因敲除的致死性。Crispr/Cas9系统也可以实现,但需依赖组织特异性Cas9递送(如AAV-TBG-Cas9),并且控制精度和可逆性远不如Cre/LoxP(但我感觉还是Crispr/Cas9好用,除非某个基因不太好靶向,只能选择全身敲,但敲了又死,长不成成年鼠,才用Loxp);
Cre/Loxp系统可以实现大段片段的重组和删除:Cre/Loxp系统可以在两个Loxp位点间的大段基因片段进行高效删除,Crispr/Cas9系统如若想实现则需要设计多个gRNA,效率低并且可能导致染色体异常;
Cre/Loxp系统可以实现基因重组和翻转:不必多言,机理如此;
复杂遗传模型构建:如果我想敲除抑癌基因同时激活原癌基因来构建癌症模型,我可以构建一个在不同基因有不同Loxp位点的小鼠,然后我只需要递送Cre酶基因载体,就可以实现两个基因的操作,但如果是Crsipr/Cas9系统,就需要涉及两个gRNA递送,比较麻烦;
可控性:两个Loxp位点交错可以实现基因的开关(虽然我感觉不太精细)。
在实际运用中,在构建复杂遗传模型时,通常是两种系统并用,以达到最大的编辑效率。
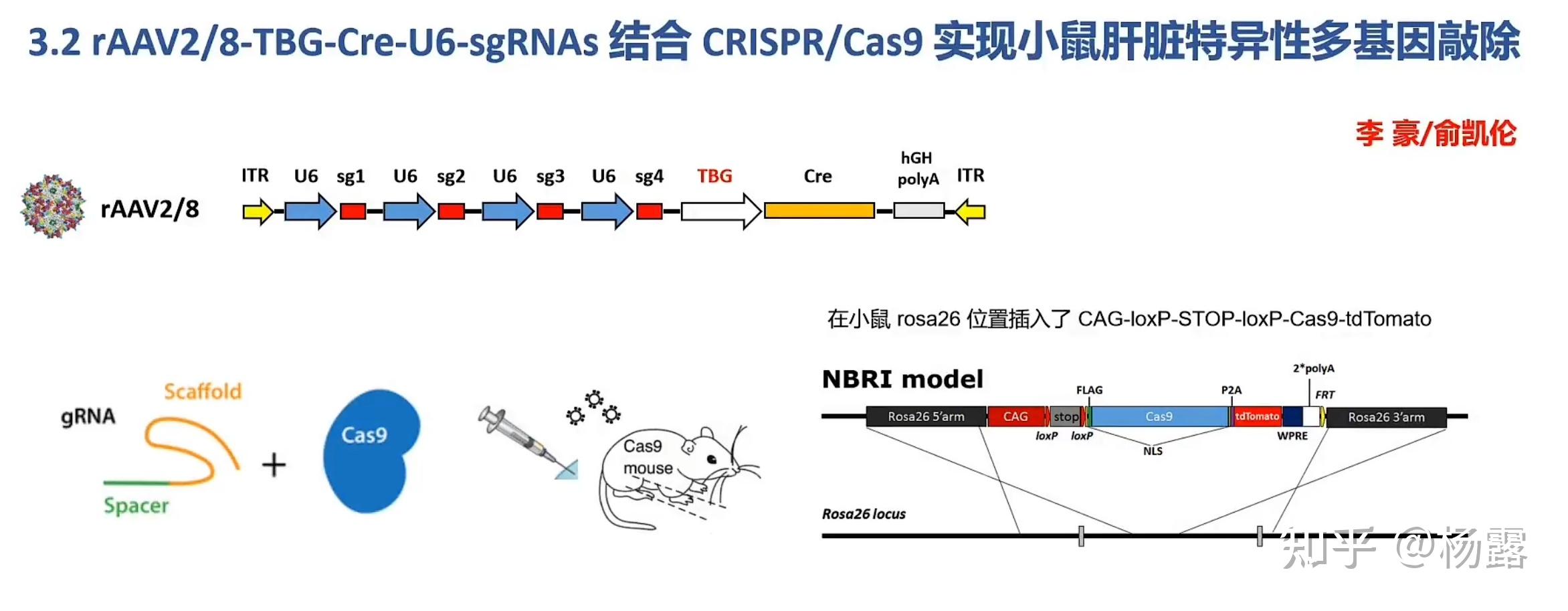
举例说明,由于Cas9酶的基因组长度远大于Cre酶基因组长度(一个是4000bp左右,一个1000多bp),因此,在构建AAV载体时,如果选用了Cas9酶基因,那么gRNA就不能太长或太多,不然包装不下;于是,一个想法诞生了:
厦大张永有教授lab
但实际上,我感觉这玩意儿不是给它搞复杂了吗?我的那个构建带有肝脏特异性启动子的Cas9基因小鼠,然后再注射肝脏靶向的载有gRNA的AAV病毒载体,双重保险,实现肝脏部位的基因特异性敲除感觉不也可以么,还简单一点。不过,我估计他这个特异性肯定比我的强,Cre系统强在特异性,不管是时空特异性还是中靶率肯定比Crispr好,但Crispr快,高效简便。
病毒载体:AAV与慢病毒
AAV详见
AAV篇章
,这里主要学习一下慢病毒载体。
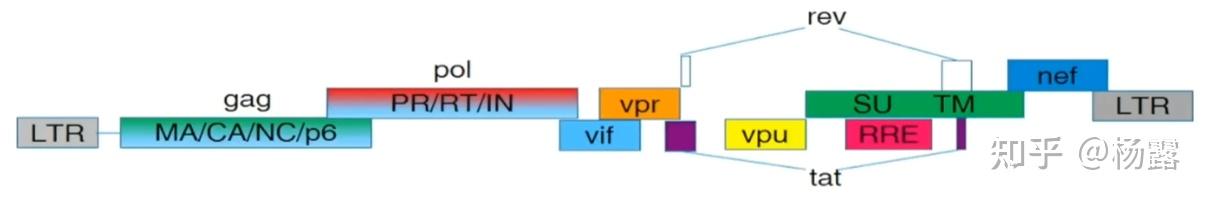
慢病毒脱胎于艾滋病病毒(HIV),经过基因修饰后去除了致病基因,保留了辅助蛋白和衣壳蛋白等基因。与AAV不同的是,慢病毒可以将目的基因整合到基因组上,实现长期且稳定的表达。
HIV基因组
LTR:长重复序列,具有一定的启动子效应,两个LTR之间的基因即是被整合到宿主细胞基因组的基因;
gag:结构核心蛋白,包括基质蛋白MA,衣壳蛋白CA,核衣壳蛋白NC等;
pol:编码核心酶的基因,包括蛋白酶PR、逆转录酶RT和整合酶IN;
env:编码病毒表面糖蛋白gp160,gp160可以剪切成为表面蛋白gp120(SU)和跨膜蛋白gp41(TM);
rev:促进病毒mRNA的核输出,并受到Rev响应元件的调控,其结合位点为RRE;
vif&vpr&vpu&nef:毒力因子,干扰宿主防御机制;
tat:激活LTR的启动子活性,启动病毒基因组转录;
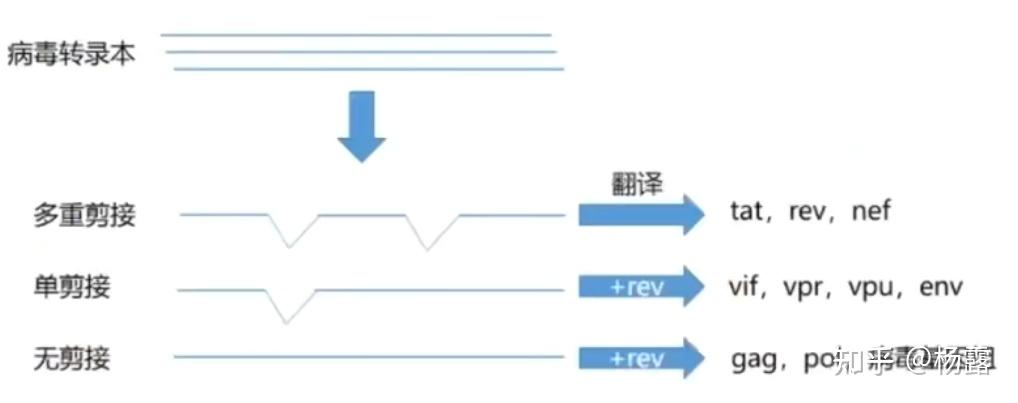
在慢病毒侵染宿主细胞时,病毒基因组整合到宿主细胞基因组,由于5'端LTR有一定的启动子活性,因此可以利用宿主细胞进行基因转录。在缺乏病毒转录激活因子tat的情况下,5'端LTR的活性很低,此时病毒基因组转录产物进行多重剪接形成短转录本,这些短转录本编码非结构蛋白tat和rev等。tat蛋白促进LTR的转录活性,促进病毒基因组所有基因的表达,rev蛋白通过结合转录本上的RRE(Rev-responsive element)来增强病毒mRNA的出核转运,并促进单剪接mRNA和无剪接mRNA的形成,其他病毒相关蛋白翻译产出,组装成新的病毒。
病毒转录过程
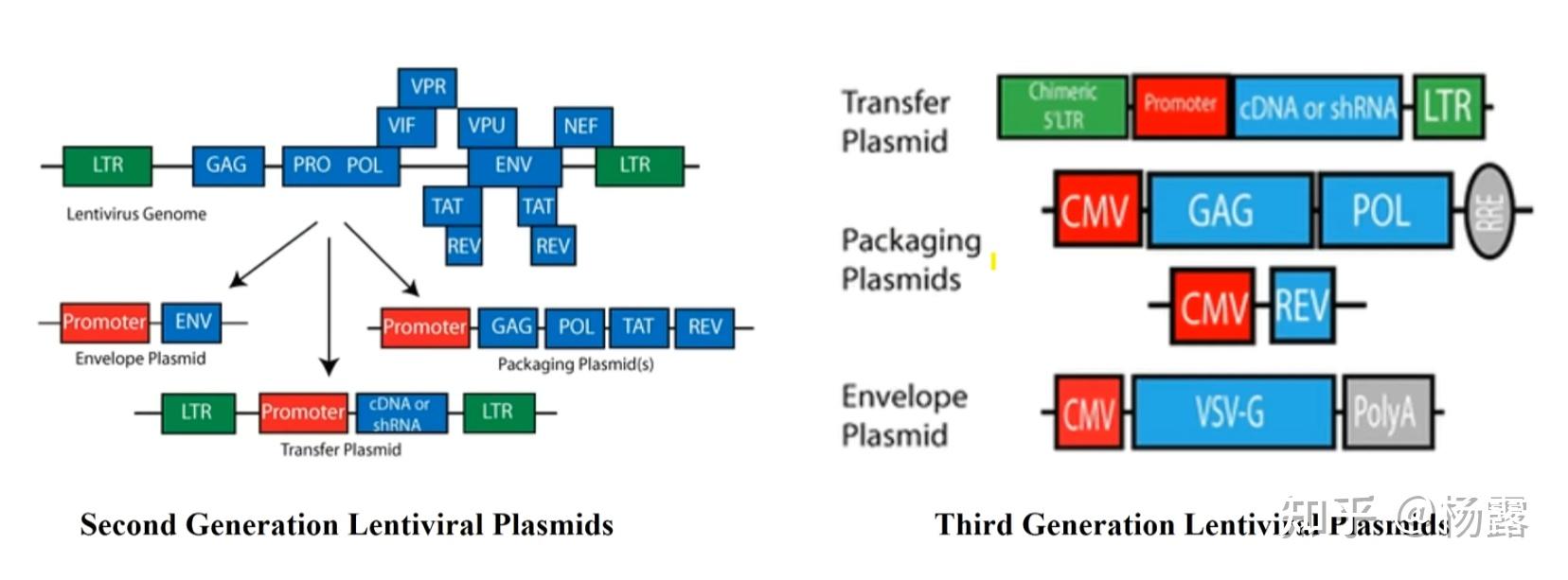
慢病毒的包装和AAV类似,经典方法也是需要三个质粒。对于二代方法而言,三个质粒,一个衣壳蛋白,一个目的基因蛋白,一个蛋白酶逆、转录酶等辅助因子。三代方法需要四个质粒,把包装质粒拆成了两个并去掉了tat,安全性更高,同时包膜糖蛋白是VSV-G,其受体更普遍。
慢病毒载体理论上在LTR之间可以插入的基因长度约为8.5 kb,但超过
3 kb
就会导致包装效率降低,并且整个质粒过大,也会导致转染效率降低,进而影响最后的滴度。
包装慢病毒载体的两代方法
相关视频和方法可以见以下资料和视频:
慢病毒和相关质粒简介
;
慢病毒包装
;
慢病毒包装详细教程
。
质粒载体
质粒是老经典的载体了,高中时就已经有接触。本来想在这里学一下质粒载体构建,但由于质粒载体构建还挺复杂的,尤其是点突变质粒载体,确实没怎么学过,因此单开一个专题:
质粒载体构建
。
如何抉择质粒载体与病毒载体:
病毒载体的典型应用
体内基因敲除(动物体):
AAV:通过局部注射(如脑、视网膜)或全身递送(如肝脏靶向AAV8)递送CRISPR组分;
慢病毒:适合需要基因组整合的长期敲除(如造血干细胞编辑);
难转染细胞:
原代T细胞、神经元等常用慢病毒或AAV转导;
质粒载体的典型应用
体外细胞系编辑:
将CRISPR质粒(含Cas9和gRNA)通过脂质体转染HEK293、HeLa等易转染细胞;
胚胎操作:
受精卵显微注射质粒DNA构建转基因小鼠(传统方法)。
总之,病毒载体适合体内实验和难转染细胞,但需权衡免疫原性和容量限制;质粒载体适合体外筛选和大片段编辑,但依赖转染效率。
原文地址:https://zhuanlan.zhihu.com/p/1895122582015035231
回复
举报
返回列表
发表回复
高级模式
B
Color
Image
Link
Quote
Code
Smilies
您需要登录后才可以回帖
登录
|
立即注册
本版积分规则
发表回复
回帖后跳转到最后一页
浏览过的版块
特定蛋白分析
关闭
官方推荐
/3
AI助手<小桔子>来了!
欢迎来交流,可以回答IVD行业各类问题!
查看 »
IVD业界薪资调查(月薪/税前)
长期活动,投票后可见结果!看看咱们这个行业个人的前景如何。请热爱行业的桔友们积极参与!
查看 »
小桔灯网视频号开通了!
扫描二维码,关注视频号!
查看 »
返回顶部
快速回复
返回列表
客服中心
搜索
洽谈合作
关注微信
微信扫一扫关注本站公众号
个人中心
个人中心
登录或注册
业务合作
-
投稿通道
-
友链申请
-
手机版
-
联系我们
-
免责声明
-
返回首页
Copyright © 2008-2024
小桔灯网
(https://www.iivd.net) 版权所有 All Rights Reserved.
免责声明: 本网不承担任何由内容提供商提供的信息所引起的争议和法律责任。
Powered by
Discuz!
X5.0 技术支持:
宇翼科技
浙ICP备18026348号-2
浙公网安备33010802005999号
快速回复
返回顶部
返回列表

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 






 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 发表于 2025-4-24 19:57
发表于 2025-4-24 19:57