金桔
金币
威望
贡献
回帖0
精华
在线时间 小时
|
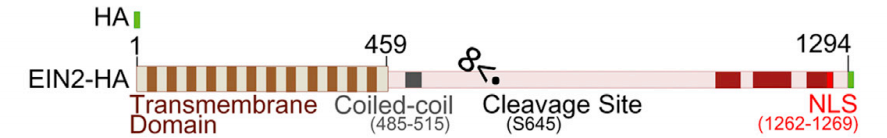
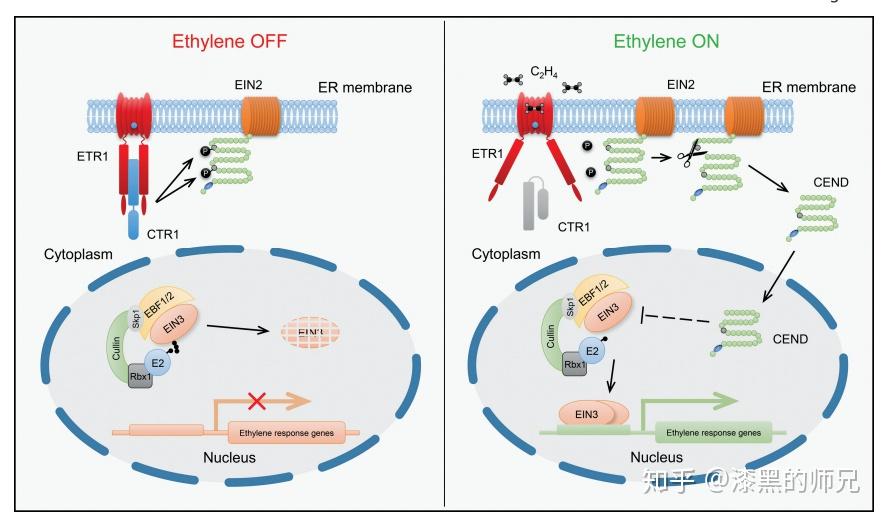
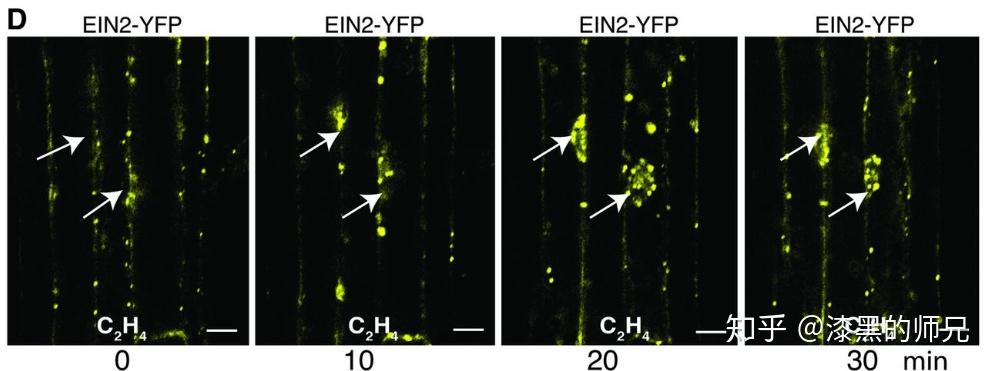
碰巧我就遇到过很多这样的蛋白。最典型的一个跟你说的一样,是一个内质网膜锚定的转录因子。既有内质网信号肽和跨膜结构,又有转录因子结构域和核定位信号。它的亚细胞定位并不唯一。
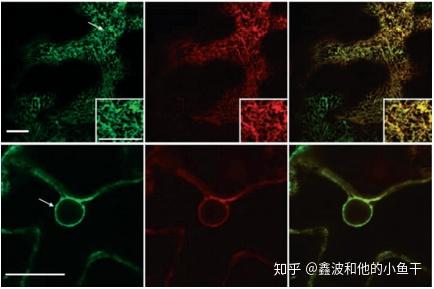
YFP标记的该蛋白(红色)与GFP标记的内质网蛋白(绿色)共定位
这个蛋白确实是一个转录因子,但是平时定位在内质网,在外界环境胁迫条件下,转录因子结构域和膜锚定区之间会被剪开(剪开它的蛋白尚未发现),掉下来的转录因子结构域会进核,发挥转录因子的功能。
一般来说,蛋白的亚细胞定位和分选信号有很大关系,分选信号的作用是引导蛋白质从胞质溶胶进入内质网、线粒体、叶绿体和过氧化物酶体,也可以引导蛋白质从细胞核进入细胞质或从高尔基体进入内质网,典型的信号序列如下。
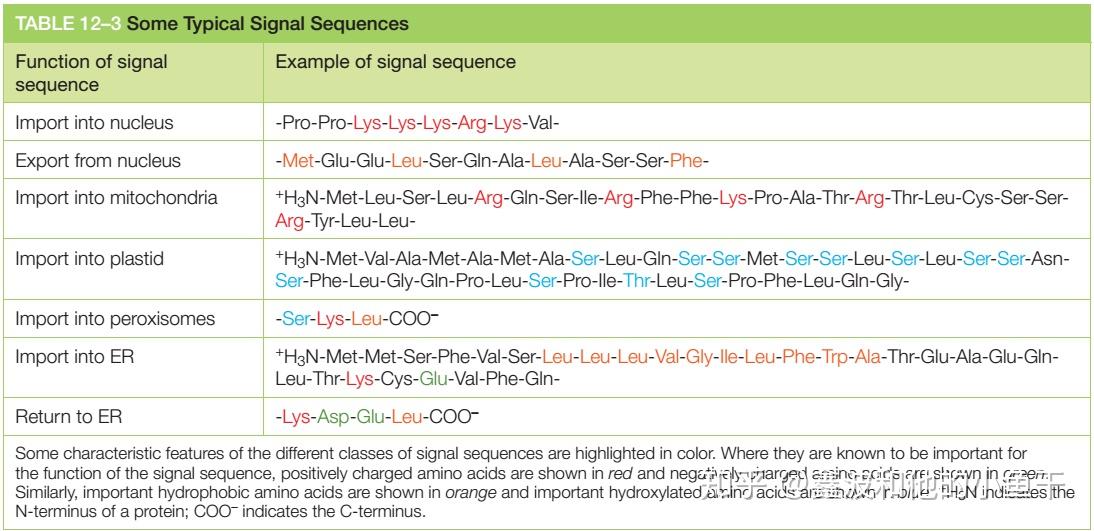
典型的信号肽序列
但是,很多蛋白不具有典型的定位信号,很多蛋白本身具有多种定位信号,很多具有典型定位信号的蛋白也不只有一种亚细胞定位。
比如有的蛋白定位在细胞膜,但是却又有极性只定位在一侧。这种情况是在太多,尤其是转运蛋白。我随便举几个例子:

水稻的Mn/Cd转运蛋白OsNramp5,定位在细胞膜,但是却显示出极性定位的特征,之定位在细胞膜的外围,接近土壤的一侧而非接近中柱的一侧,作用于Mn的吸收。
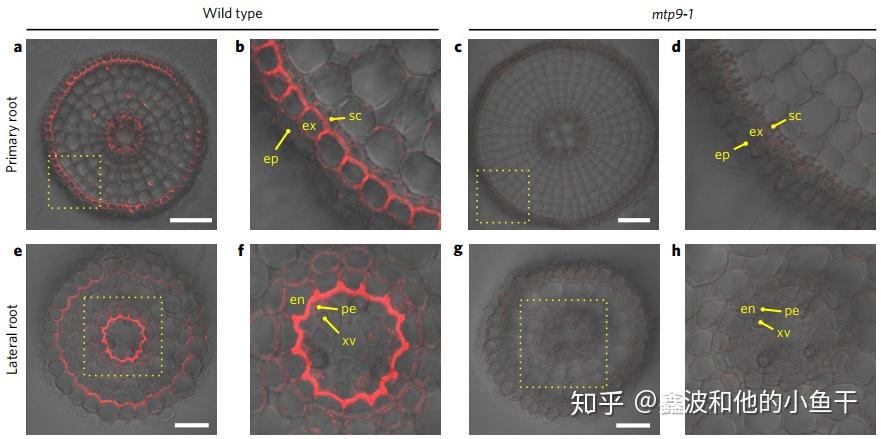
MTP9与Nramp5相反,刚好定位在中柱一侧,作用于Mn的木质部装载
与OsNramp5和OsMTP9类似的还有作用于Si吸收和转运的通道蛋白Lsi1和Lsi2.
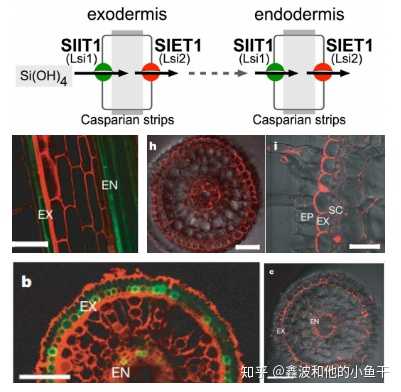
Lsi1与Lsi2的亚细胞定位
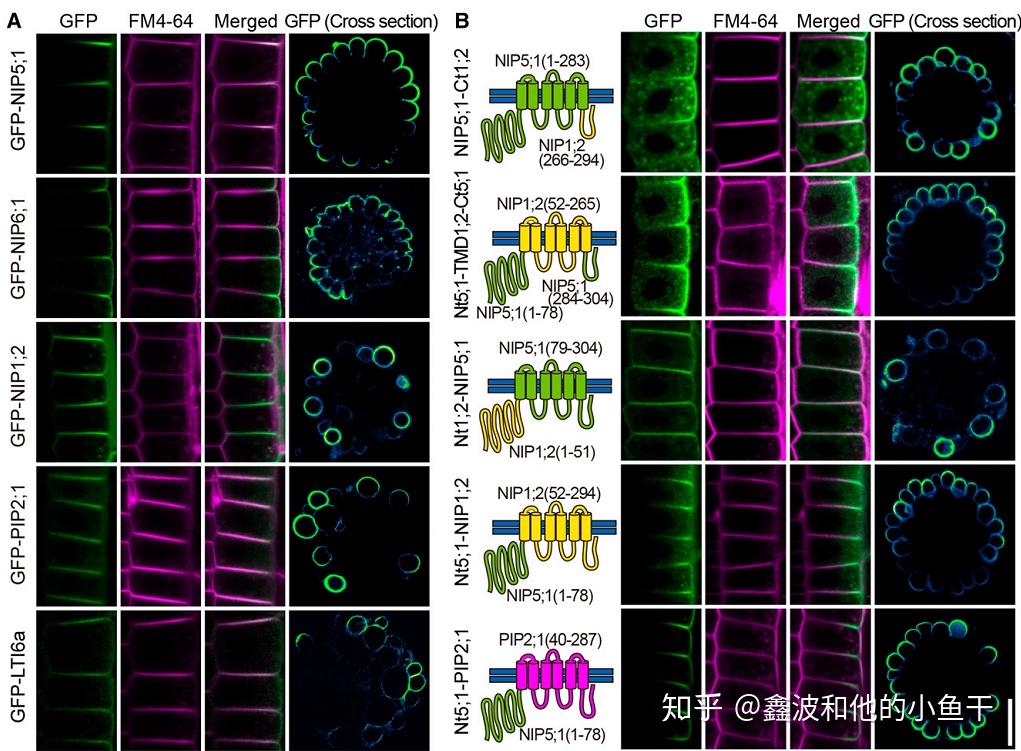
拟南芥的水通道蛋白Nip5;1和Nip6;1定位在细胞外围,极性定位的特征是由N端决定的,N端的TPGTPGTPG序列中T被磷酸化后显示出极性定位,去除N端或者将T突变成A阻止其磷酸化则失去极性定位能力,将T突变成D模拟磷酸化则表现为组成型的极性定位。
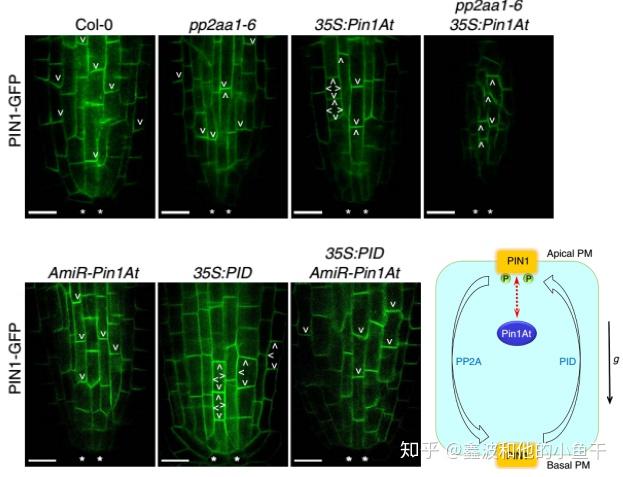
尽管生长素转运蛋白PIN1定位在细胞膜,但是并不是细胞膜的每一侧都有,PID和PP2A通过可逆磷酸化影响生长素转运蛋白Pin1,Pin1At识别并对其变构,影响它的极性定位。
除了转运蛋白的亚细胞定位,还有很多转录因子、光受体等蛋白的亚细胞定位不唯一,在接受到外界信号后会发生改变,到该去的地方发挥它的功能,都是动态的过程不是死的。甚至糖酵解里边的酶也会进入细胞核,作为核内受体发挥功能。我随便举几个例子:
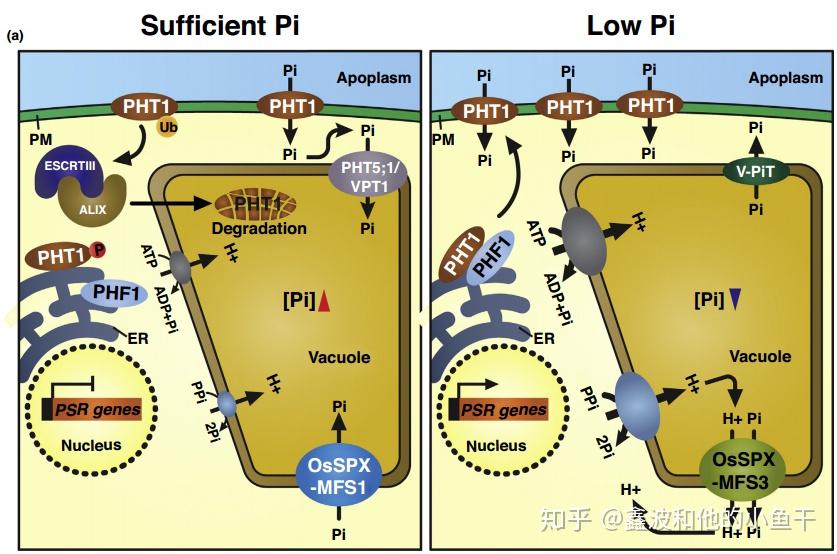
Pi充足的情况下PHT1被磷酸化,滞留在内质网,而缺磷的情况下,PHT1去磷酸化并被PHF1带到细胞膜。
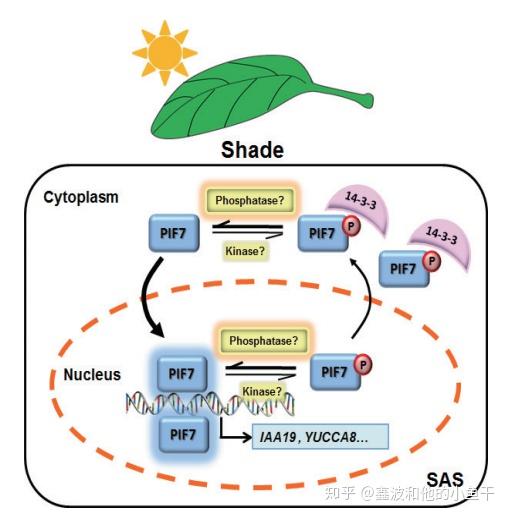
接收到光信号后,转录因子PIF7被激酶磷酸化激活进核,发挥功能后被磷酸酶去磷酸化出核,14-3-3蛋白识别磷酸化的PIF7并参与其进出核的过程。
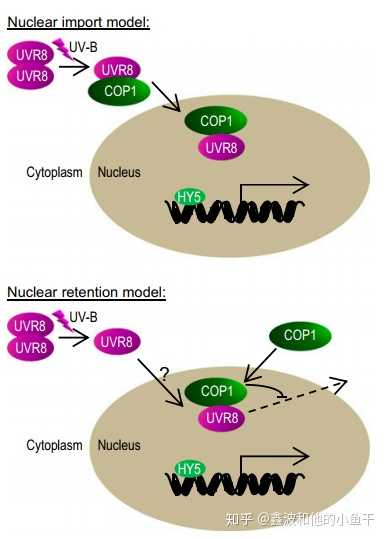
紫外线受体UVR8定位于胞质和细胞核,但其发挥主要活性被认为是在细胞核。UV-B光感受刺激UVR8的核积聚。COP1在调控UV-B诱导的UVR8核积聚和UVR8介导的紫外光信号转导中具有双重作用。
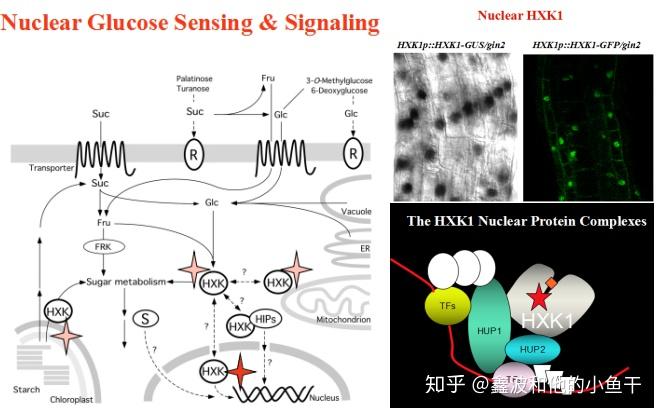
HXK1作为一个传统的糖酵解酶,不单单是存在在细胞质,线粒体和叶绿体中的。它也有很小的一部分定位在细胞核中。在高光照条件下,可以清楚的发现HXK1的核定位。作为葡萄糖的受体,HXK1可以感受糖的信号,进入细胞核,结合HUP1,HUP2, 然后这个复合体包括一些转录因子一起调控葡萄糖相关基因的表达。
更多关于蛋白分选的内容可移步我的专栏,查阅这些文章(更新中):鑫波和他的小鱼干:细胞内功能区隔与蛋白质分选,鑫波和他的小鱼干:内质网和高尔基体的膜泡运输——膜泡的形成。
参考文献
- Sasaki A , Yamaji N , Yokosho K , et al. Nramp5 Is a Major Transporter Responsible for Manganese and Cadmium Uptake in Rice[J]. The Plant Cell, 2012, 24(5):2155-2167.
- Ueno D , Sasaki A , Yamaji N , et al. A polarly localized transporter for efficient manganese uptake in rice[J]. Nature Plants, 2015, 1(12):15170.
- Ma J F , Tamai K , Yamaji N , et al. A silicon transporter in rice[J]. Nature, 2006, 440(7084):688-691.
- Wang S , Yoshinari A , Shimada T , et al. Polar Localization of the NIP5;1 Boric Acid Channel Is Maintained by Endocytosis and Facilitates Boron Transport in Arabidopsis Roots[J]. The Plant Cell, 2017:tpc.00825.2016.
- Xu H , Qian Z , Yupei J , et al. Shade-induced nuclear localization of PIF7 is regulated by phosphorylation and 14-3-3 proteins in Arabidopsis[J]. eLife, 2018, 7:e31636-.
- Yin R , Skvortsova M Y , Loubéry, Sylvain, et al. COP1 is required for UV-B–induced nuclear accumulation of the UVR8 photoreceptor[J]. Proceedings of the National Academy of Sciences, 2016:201607074.
- Moore, B. Role of the Arabidopsis Glucose Sensor HXK1 in Nutrient, Light, and Hormonal Signaling[J]. Science, 2003, 300(5617):332-336.
|
|

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 






 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 发表于 2025-1-29 13:06
发表于 2025-1-29 13:06