用户名
UID
Email
密码
记住
立即注册
找回密码
只需一步,快速开始
微信扫一扫,快速登录
开启辅助访问
快捷导航
门户
Portal
社区
BBS
资讯
会议
市场
产品
问答
数据
专题
帮助
签到
每日签到
企业联盟
人才基地
独立实验室
产业园区
投资机构
检验科
招标动态
供给发布
同行交流
悬赏任务
共享资源
VIP资源
百科词条
互动话题
导读
动态
广播
淘贴
法规政策
市场营销
创业投资
会议信息
企业新闻
新品介绍
体系交流
注册交流
临床交流
同行交流
技术杂谈
检验杂谈
今日桔说
共享资源
VIP专区
企业联盟
投资机构
产业园区
业务合作
投稿通道
升级会员
联系我们
搜索
搜索
本版
文章
帖子
用户
小桔灯网
»
社区
›
C、IVD技术区
›
原料技术
›
酶活性的调节方式(一)
图文播报
2026庆【网站十三周
2025庆【网站十二周
2024庆中秋、迎国庆
2024庆【网站十一周
2023庆【网站十周年
2022庆【网站九周年
返回列表
查看:
10362
|
回复:
0
[分享]
酶活性的调节方式(一)
[复制链接]
Rose
Rose
当前离线
金桔
金币
威望
贡献
回帖
0
精华
在线时间
小时
发表于 2024-11-11 17:46
|
显示全部楼层
|
阅读模式
登陆有奖并可浏览互动!
您需要
登录
才可以下载或查看,没有账号?
立即注册
×
酶的活性调节主要包括四种方式:变构调节、共价修饰调节、酶原激活以及调节蛋白的调控。
有些酶属于别构蛋白,即在专一性效应物的诱导下,结构发生变化,使催化活性改变,称为变构酶或别构酶(allosteric enzyme)。使酶活增加的效应物称为正调节物,反之称为负调节物。变构酶都是寡聚酶,分子中除活性中心外还有别构中心(调节中心)。两个中心可在同一亚基,也可在不同亚基。有活性中心的亚基称为催化亚基,有别构中心的亚基称为调节亚基。
别构酶还有同促效应和异促效应,前者是指酶的底物同时可以作为别构调节物,后者就是指底物和调节物是不同的分子。
大部分别构酶的v-[S]曲线呈S形,与米氏酶不同。这种曲线表明酶与一分子底物(或效应物)分子结合后,其构象发生改变,有利于后续分子的结合,称为正协同效应。这种现象有利于对反应速度的调节,在未达到最大反应速度时,底物浓度的略微增加,将使反应速度有极大提高。所以正协同效应使酶对底物浓度的变化极为敏感。
另一类别构酶具有负协同效应,其动力学曲线类似双曲线,在底物浓度极低时反应速度变化很快,但继续下去则速度变化缓慢。所以负协同效应使酶对底物浓度变化不敏感。
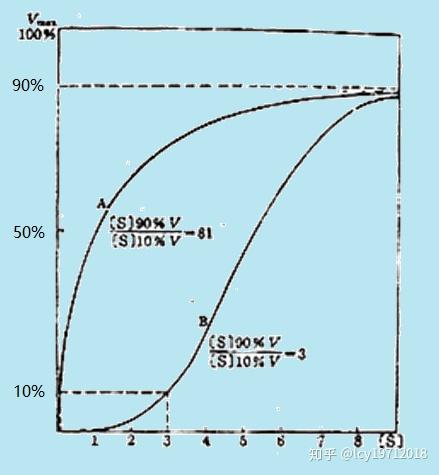
因为一些没有别构效应的酶也可产生类似的v-[S]曲线,所以作图法不能完全作为判断别构酶的依据。可用Rs值(saturation ratio,饱和比值)([S]90%V / [S]10%V)来定量地区分三种酶:Rs=81为米氏酶,小于81为正协同效应,大于81为负协同。还可采用Hill系数法,以lg(v/(Vm-v))对lg[S]作图,曲线的最大斜率h称为Hill系数,米氏酶等于1,正协同酶大于1,负协同小于1。
米氏酶与正协同酶的Rs值
关于别构酶的变构机制,有两种模型。齐变模型(M. W. C.)认为酶分子中所有亚基的构象相同,无杂合状态。在低活性的紧张态(tight,T)和高活性的松弛态(relaxed form,R)之间存在平衡,效应物使平衡移动,从而改变酶的活性。此模型不适于负协同的酶。
序变模型(K.N.F.)则认为各个亚基可以杂合存在,变构是由于配体的诱导,而不是因为平衡的移动。协同性取决于与配体结合的亚基对空位亚基的影响。此模型对两种酶都适用。
天冬氨酸转氨甲酰酶(ATCase)是嘧啶合成途径的第一个酶,共12个亚基,其中催化和调节亚基各6个。分子结构为2个C3中间夹着3个R2,活性中心位于两个催化亚基中间。别构中心位于调节亚基的远端,通过变构影响催化亚基的活性。
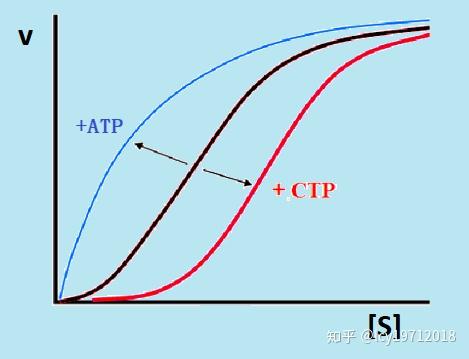
此酶受到CTP的反馈抑制,可被ATP激活。Asp、氨甲酰磷酸均有正同促效应,CTP有异促效应,可使酶的S形程度增大,即Rs值减小,CTP之间具有正协同作用,n=3。ATP使Rs增大,当达到饱和时即成为双曲线。ATP和CTP都只改变酶的亲和力,而不影响Vm。琥珀酸是天冬氨酸的类似物,在天冬氨酸浓度高时是竞争性抑制剂,而当天冬氨酸不足时则可模拟天冬氨酸的正调控变构作用而成为激活剂。
ATCase的别构调节
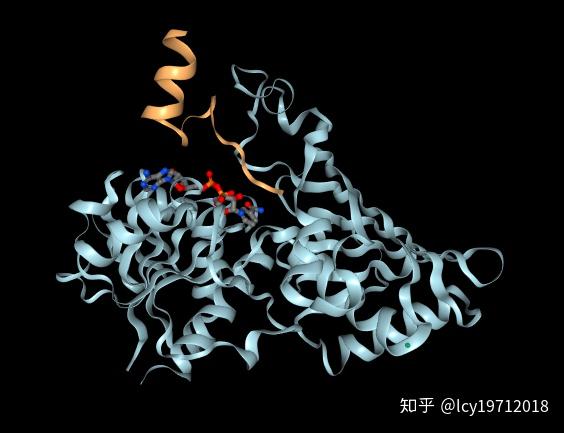
磷酸甘油醛脱氢酶(GAPDH)是负协同酶的典型代表。共四个亚基,Km1和Km2都较小,易与NAD+结合,即在低底物浓度时反应较快;而Km3则增大了100倍,很难与NAD+反应。这是由构象变化引起的。在生物体内,当NAD+不足时可以保证酵解的进行,而当过NAD+多时则供给其它反应,避免造成酸中毒。
GAPDH的一个亚基
留个思考题:别构抑制与非竞争抑制是否相同?更进一步,别构抑制剂算不算抑制剂?
原文地址:https://zhuanlan.zhihu.com/p/72432778
回复
举报
返回列表
发表回复
高级模式
B
Color
Image
Link
Quote
Code
Smilies
您需要登录后才可以回帖
登录
|
立即注册
本版积分规则
发表回复
回帖后跳转到最后一页
浏览过的版块
化学发光技术
检验资讯
质谱技术
招标动态
科学研究
关闭
官方推荐
/3
AI助手<小桔子>来了!
欢迎来交流,可以回答IVD行业各类问题!
查看 »
IVD业界薪资调查(月薪/税前)
长期活动,投票后可见结果!看看咱们这个行业个人的前景如何。请热爱行业的桔友们积极参与!
查看 »
小桔灯网视频号开通了!
扫描二维码,关注视频号!
查看 »
返回顶部
快速回复
返回列表
客服中心
搜索
洽谈合作
关注微信
微信扫一扫关注本站公众号
个人中心
个人中心
登录或注册
业务合作
-
投稿通道
-
友链申请
-
手机版
-
联系我们
-
免责声明
-
返回首页
Copyright © 2008-2024
小桔灯网
(https://www.iivd.net) 版权所有 All Rights Reserved.
免责声明: 本网不承担任何由内容提供商提供的信息所引起的争议和法律责任。
Powered by
Discuz!
X5.0 技术支持:
宇翼科技
浙ICP备18026348号-2
浙公网安备33010802005999号
快速回复
返回顶部
返回列表

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 






 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 发表于 2024-11-11 17:46
发表于 2024-11-11 17:46