用户名
UID
Email
密码
记住
立即注册
找回密码
只需一步,快速开始
微信扫一扫,快速登录
开启辅助访问
快捷导航
门户
Portal
社区
BBS
资讯
会议
市场
产品
问答
数据
专题
帮助
签到
每日签到
企业联盟
人才基地
独立实验室
产业园区
投资机构
检验科
招标动态
供给发布
同行交流
悬赏任务
共享资源
VIP资源
百科词条
互动话题
导读
动态
广播
淘贴
法规政策
市场营销
创业投资
会议信息
企业新闻
新品介绍
体系交流
注册交流
临床交流
同行交流
技术杂谈
检验杂谈
今日桔说
共享资源
VIP专区
企业联盟
投资机构
产业园区
业务合作
投稿通道
升级会员
联系我们
搜索
搜索
本版
文章
帖子
用户
小桔灯网
»
社区
›
D、仪器区
›
免疫仪器
›
Toll样受体:发现免疫探测器的起点(下) ...
图文播报
2026庆【网站十三周
2025庆【网站十二周
2024庆中秋、迎国庆
2024庆【网站十一周
2023庆【网站十周年
2022庆【网站九周年
返回列表
查看:
7531
|
回复:
0
[分享]
Toll样受体:发现免疫探测器的起点(下)
[复制链接]
营养师
营养师
当前离线
金桔
金币
威望
贡献
回帖
0
精华
在线时间
小时
发表于 2024-9-24 11:45
|
显示全部楼层
|
阅读模式
<
登陆有奖并可浏览互动!
您需要
登录
才可以下载或查看,没有账号?
立即注册
×
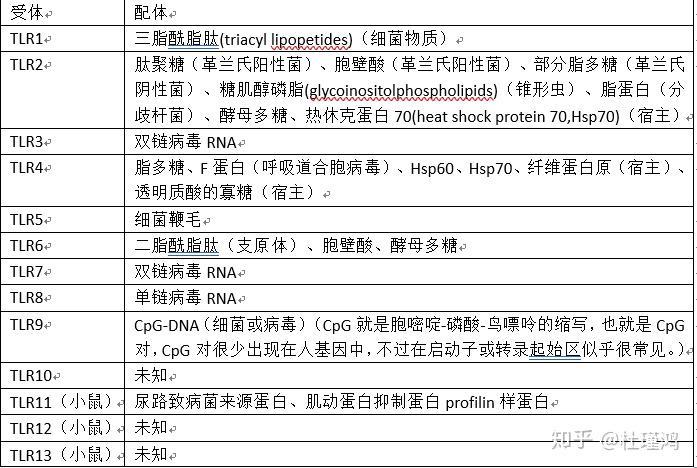
人们陆续发现了13种TLR,也就将它们分别命名为TLR1-13。它们能识别的生物物质可以见于下表:
读者可以看到TLR能够识别的化学物质既有来自细菌和病毒的,也有来自自身(宿主)的。实际上,内源性的配体一般在机体应激或是组织损伤时释放。
在配体与TLR结合以后,LTR就会两两聚在一起,空间结构随之发生变化。在这之后,就是复杂的级联通讯反应。其中的一条路径是,TLR二聚以后结合髓样分化因子88(Myeloid differentiation factor 88,MyD88)。Toll样受体是一个跨膜的蛋白质,它有一部分位于细胞外,用来识别自己的配体;有一部分位于细胞内,这部分包含了一个特殊的部位(区域),叫作“Toll/IL-1受体同源区”(Toll/IL-1R homologous region,TIR)。TIR就是LTR与MyD88结合的地方。MyD88这种蛋白质的游离羧基端有一个TIR结构域,游离氨基端有一个“死亡结构域”。在配体还没与TLR结合时,MyD88与β-肌动蛋白结合在一起,而后者正是细胞骨架的一部分,所以MyD88那个时候被锚定在细胞骨架上;此后将会与MyD88结合的白介素-1受体相关激酶1(interleukin-1 receptor-associated kinase,IRAK1),在还没受到配体的刺激时,则与MyD88调节蛋白-Toll互作蛋白(Toll interacting protein,Tollip)结合在一起。配体刺激细胞导致细胞骨架重排,MyD88便被释放,并向着TLR的TIR聚集;IRAK1被MyD88的死亡结构域募集的IRAK4结合并被后者磷酸化(这个过程也许有另外两种蛋白质——MyD88适配体样蛋白(MyD88 adaptor-like,MAL)和布鲁顿酪氨酸蛋白激酶(Bruton Tyrosine-protein kinase,BTK)的协助),导致它从Tollip那被释放出来,去与MyD88的死亡结构域结合,这样一来,IRAK1被激活,它可以把自己再磷酸化一次,导致自己与MyD88分离,并募集和结合肿瘤坏死因子受体活化因子6(tumor necrosis factor(TNF) receptor activating factor 6,TRAF6)并将它激活,激活后的TRAF6又与转化生长因子-β活化激酶1(transforming growth factor-β activating kinase,TAK1)和TAK结合蛋白——TAB1和TAB2形成复合体。这个复合体具有很多功能,比如激活NEMO(NF-κB-essential modulator,又叫IKKγ)-IKK1(IκB kinase 1,即IκB激酶1)-IKK2复合体,这个复合体会结合核因子κB抑制蛋白(IκB),将后者磷酸化。于是乎本来与NF-κB结合着,导致它被抑制的IκB脱离了NF-κB,对NF-κB的抑制就被解除,NF-κB的浓度就增加,它就进入细胞核——而通过我们往期的介绍,你应该已经知道了NF-κB是个功能强大的转录因子。此外,如果在TRAF6与TAK1、TAB1和TAB2形成复合蛋白质时,还可能结合另外一个叫作“ECSIT”(全称是Evolutionarily conserved signaling intermediate in Toll pathway(进化保守的在Toll通路中的信号中介(蛋白)))的蛋白质,这个操作可以将TRAF6与其他的级联通讯途径联系起来,然后激活更多基因的表达。此外它还可以激活其他各种各样的通路,所有这些通路的最终效应是激活一揽子基因的表达。这个路径就是所谓的MyD88依赖性通路。除了上面介绍的这个级联通讯路径以外,MyD88依赖性通路还有一个途径,那就是MyD88→TRAF3→IRAF3→TBK1(TRAF-family-member-associated NF-κB activator-binding kinase,即TRAF家族成员相关的NF-κB激活蛋白结合激酶)→IRF7(IFN regulatory factor 7,即干扰素调节因子7),这样前面一个激活后面一个,直到IRF7,IRF7再去刺激干扰素α的表达。
还有一个MyD88非依赖性通路,就是说配体在结合TLR以后不需要MyD88的参与。那么一开始充当结合TLR的TIR的蛋白质,可能是TRAM(TIR domain-containing adaptor molecule,即包含TIR结构域的适配分子)等等。TRIF能够作用于两种蛋白质——IKKε-TBK1(TBK1就是端锚蛋白结合激酶1,在干扰素一节中,我们曾略有提到)和受体互作蛋白1(receptor-interacting protein 1)。IKKε-TBK1能激活IRF3(就是干扰素调节因子3),IRF3再进入细胞核去刺激干扰素刺激反应元件(interferon-stimulated response element,ISRE),从而调节干扰素的表达;而RIP1则会去刺激NEMO-IKK1-IKK2磷酸化IκB,然后激活NF-κB。
上面说到的这些级联通讯网络,并不是所有的LTR都会用到。比如,LTR3不会结合MyD88,因此也就没有MyD88依赖性通路。而LTR4两种通路都可以用,其他的LTR只能使用MyD88依赖性通路来刺激基因表达。经由TLR引发的基因表达的结果,是一系列的免疫活动和炎症反应。比如,促进内皮细胞表达更多的选择素,从而让更多白细胞离开血管和脉管并跟踪甚至攻击入侵的病原体;激活巨噬细胞和中性粒细胞并增强它们的吞噬能力;也许还能让一些免疫细胞产生诸如一氧化氮这样的抗菌物质;激活自然杀伤细胞、单核细胞、嗜碱性粒细胞、嗜酸性粒细胞等白细胞,促使它们释放炎症因子、趋化因子;间接地激活T细胞和B细胞从而引发获得性免疫反应……有的TLR反应也许还与自身免疫病有关。炎性反应具有清除致病因子和坏死组织(引发白细胞聚集和体温升高,一定程度的体温升高能够使机体代谢增强,促进抗体形成与释放,增加吞噬细胞的吞噬能力)、稀释毒素(导致充血、血管扩张、血管通透性增强和血液成分渗出与水肿)和引发组织增生的作用。当然,也可能如果机体受到了过多的刺激,就会引发过于强烈的炎症反应——比如,过于强烈的巨噬细胞甚至会转变为“炎性嗜血细胞”,攻击并吞噬红细胞和其他的自身组织。但是,正如读者所见,一旦牵扯到干扰素、趋化因子、细胞因子和炎症那些事儿,我们就讲不清楚了。先天免疫系统十分复杂,人们对它们的工作方式认识得可没有那么明白。因此,如今在免疫学的研究中,一门新兴的学科“炎症学”似乎正在慢慢形成。在人们探索这个领域的道路上出现的每一条骇人听闻的新闻——就比如“炎性嗜血细胞”和嗜血细胞综合征的发现——都能增进人们对炎症和免疫系统的理解。
原文地址:https://zhuanlan.zhihu.com/p/93456950
回复
举报
返回列表
发表回复
高级模式
B
Color
Image
Link
Quote
Code
Smilies
您需要登录后才可以回帖
登录
|
立即注册
本版积分规则
发表回复
回帖后跳转到最后一页
浏览过的版块
凝血仪器
尿液分析仪器
生物安全
核酸提取
室间质评
关闭
官方推荐
/3
AI助手<小桔子>来了!
欢迎来交流,可以回答IVD行业各类问题!
查看 »
IVD业界薪资调查(月薪/税前)
长期活动,投票后可见结果!看看咱们这个行业个人的前景如何。请热爱行业的桔友们积极参与!
查看 »
小桔灯网视频号开通了!
扫描二维码,关注视频号!
查看 »
返回顶部
快速回复
返回列表
客服中心
搜索
洽谈合作
关注微信
微信扫一扫关注本站公众号
个人中心
个人中心
登录或注册
业务合作
-
投稿通道
-
友链申请
-
手机版
-
联系我们
-
免责声明
-
返回首页
Copyright © 2008-2024
小桔灯网
(https://www.iivd.net) 版权所有 All Rights Reserved.
免责声明: 本网不承担任何由内容提供商提供的信息所引起的争议和法律责任。
Powered by
Discuz!
X5.0 技术支持:
宇翼科技
浙ICP备18026348号-2
浙公网安备33010802005999号
快速回复
返回顶部
返回列表

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 






 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 发表于 2024-9-24 11:45
发表于 2024-9-24 11:45