用户名
UID
Email
密码
记住
立即注册
找回密码
只需一步,快速开始
微信扫一扫,快速登录
开启辅助访问
快捷导航
门户
Portal
社区
BBS
资讯
会议
市场
产品
问答
数据
专题
帮助
签到
每日签到
企业联盟
人才基地
独立实验室
产业园区
投资机构
检验科
招标动态
供给发布
同行交流
悬赏任务
共享资源
VIP资源
百科词条
互动话题
导读
动态
广播
淘贴
法规政策
市场营销
创业投资
会议信息
企业新闻
新品介绍
体系交流
注册交流
临床交流
同行交流
技术杂谈
检验杂谈
今日桔说
共享资源
VIP专区
企业联盟
投资机构
产业园区
业务合作
投稿通道
升级会员
联系我们
搜索
搜索
本版
文章
帖子
用户
小桔灯网
»
社区
›
C、IVD技术区
›
PCR技术
›
国家首席科学家石磊教授发明的“恒温PCR技术”是怎么样 ...
图文播报
2026庆【网站十三周
2025庆【网站十二周
2024庆中秋、迎国庆
2024庆【网站十一周
2023庆【网站十周年
2022庆【网站九周年
返回列表
查看:
11086
|
回复:
3
[分享]
国家首席科学家石磊教授发明的“恒温PCR技术”是怎么样的一种技术?
[复制链接]
虎威将军
虎威将军
当前离线
金桔
金币
威望
贡献
回帖
0
精华
在线时间
小时
发表于 2024-9-21 10:26
|
显示全部楼层
|
阅读模式
登陆有奖并可浏览互动!
您需要
登录
才可以下载或查看,没有账号?
立即注册
×
这种技术在国外是否就已经有类似的技术基础参考?它是怎样打破传统不用通过常规pcr的调节温度来扩增的?在线等,急希望各位大神帮帮忙解答一下。
原文地址:https://www.zhihu.com/question/27269423
回复
举报
队长是我
队长是我
当前离线
金桔
金币
威望
贡献
回帖
0
精华
在线时间
小时
发表于 2024-9-21 10:27
|
显示全部楼层
对于喜欢恒温扩增的朋友来说,这篇文章值得一读,它的原文:
Highly specific, multiplexed isothermal pathogen detection with fluorescent aptamer readout - PMC (nih.gov)
,有余力的可以看看。
上海前迪生物科技 上海前迪生物科技有限公司
2023-09-16 22:44
发表于上海
一摘要:
采用等温、无细胞、合成生物学方法进行病原体检测,是利用生物系统中一些放大的能力,例如不用复杂的活细胞系统,而以冻干形式存在的聚合酶依然保持着酶的高活性,其中基于核酸序列的扩增(NASBA)是其中最为出名的。尽管与细胞系统相比降低了复杂性,但这些系统中的副反应还是经常发生的。因此,这些系统经常会表现出假阳性。在我们的试验中,我们证明包含缺乏启动子且与扩增产物无关的DNA双链可以完全抑制假阳性。标记合适的荧光适配体做为NASBA标签(Apta-NASBA)。Apta-NASBA具有1 pM的检测限,并且可以提供多重、多色荧光检测。此外,Apta-NASBA可以使用多种设备进行检测,例如荧光微孔板读数仪、qPCR仪器或基于Raspberry Pi的3D打印检测平台(使用手机相机模块)兼容实地检测。
二、介绍:
低成本、快速的病原体检测方法对现场诊断在发展中国家用于疾病检测和病原体监测(例如,在供水或农产品中)都非常适用并有较大的应用应用范围。目前已经开发了几种核酸扩增技术来鉴定致病病原体,其中聚合酶链式反应(PCR)是最著名的方法(Garibyan和Avashia,2013)。聚合酶链式反应具有强大、特异性和广泛应用性,是广泛诊断测试的基础。然而,进行PCR所需的热循环器是需要大量能源且昂贵的(~5–10K美元),这使得它们在现场诊断或缺乏能源的地方中使用的成本过高。此外,PCR的典型检测,如用电泳电泳方法进行检测,需要经过培训的技术人员和额外的设备。实时监测PCR反应是可能的,但这需要荧光检测系统与热循环仪的耦合,从而使仪器的成本增加了一个数量级。这些反应通常通过昂贵的寡核苷酸探针或非特异性DNA嵌入染料来监测(Arya等人,2005)。
等温核酸扩增技术是降低与热循环相关的成本和复杂性的一种有吸引力的手段。已经报道了几种等温技术,如基于核酸序列的扩增(NASBA)、环介导的扩增(LAMP)、重组酶聚合酶扩增(RPA)、解旋酶依赖性扩增和SHERLOCK(Notomi等人2000;Deiman等人2002;An等人2005;Piepenburg等人2006;Gootenberg等人2017)。在所有这些反应中,它们使用链置换、核酸酶或重组酶来生成双链DNA,或者生成作为产物或转录为RNA输出的中间体的双链DNA。产生RNA输出的反应受到高度关注,因为多种RNA聚合酶可以容易地过度表达、冻干(不需要冷链)并用于在这些反应中生成大量RNA。此外,与双链DNA不同,RNA可以表现出多种功能行为,包括荧光染料的结合和酶的活性,从而实现多种潜在的检测方法(Abdolahzadeh等人,2019)。
与等温扩增反应相关的一个常见缺陷是非特异性扩增,这不能像常规PCR一样通过热循环来缓解。研究人员使用了一系列技术来缓解这一缺陷,包括工程化的热稳定聚合酶和针对目标扩增子不同区域的两步嵌套反应(包括另一种基于荧光适体的方法,nested Mango NASBA)。这些策略虽然具有高灵敏度和特异性,但必然会增加反应的复杂性(Abdolahzadeh等人,2019;吴等人2020)。
这段文字讨论了在反应中使用了病毒聚合酶(例如T7 RNA聚合酶)的非特异性扩增的一个潜在来源,即不需要启动子或增强子等调控元件,便可以直接进行转录的过程。最近发现,T7 RNA聚合酶的这种潜在行为是该酶产生的转录物表现出先天免疫激活的原因。这是因为该酶产生一种低水平的反义转录物,与主要的正义转录物杂交产生双链RNA(dsRNA)并导致先天免疫激活(Mu等人,2018)。作者推测,未被促进的转录可能在NASBA反应中起作用,产生一个编码荧光适体的引物二聚体,该二聚体可以继续参与反应。在这里,作者证明了加入一个不包含启动子序列的竞争二聚体可以完全抑制NASBA的非特异性扩增。使用这种技术,作者展示了用多种不同序列和结构的荧光适体对NASBA扩增进行多色标记,包括在多重、单管反应中进行实时检测。
三、结果与讨论:
Apta-NABSA涉及使用两个引物:第一个是常规的NASBA引物,它安装T7RNA聚合酶启动子序列。第二个引物结合扩增子,并另外包含功能性RNA的反义序列(这里是荧光适配体)。因此,随着RNA扩增子转录的数量增加,RNA适配体的数量也增加。然后适配体可以折叠并与其相应的小分子结合,产生特定的、实时的荧光信号。使用编码荧光适配体的引物尤其吸引人,因为它可以将通常与双重标记探针(例如TaqMan、分子信标)相关的特异性,与小分子染料(例如SYBRGreen)的低成本相结合。我们使用以前为实时PCR开发的引物,设计了针对肠聚集性大肠杆菌毒力基因aggR的引物,aggR编码聚集性粘附菌毛I的转录激活剂。缺乏T7RNA聚合酶启动子序列的引物包含Broccoli适体的编码序列,当与荧光染料DFHBI-1T结合时,可作为基因编码的绿色荧光RNA标记。
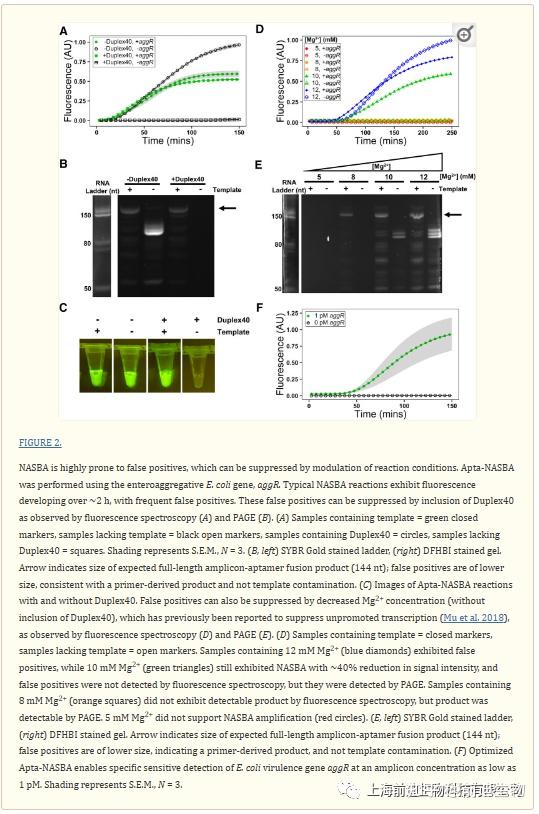
正如预期的那样,含荧光适体的引物编码实现了指数级扩增曲线(图2A)。然而,正如在NASBA反应中所观察到的那样,在不含模板的对照中也发生了非模板介导的扩增(图2A-E;Abdolahzadeh等,2019)。T7 RNA聚合酶是众所周知的,其催化范围远超其经典的启动子驱动的DNA依赖性RNA聚合酶活性,包括RNA依赖性RNA聚合酶活性(Konarska和Sharp,1989)和非启动子依赖的DNA依赖性聚合,如最近报道的(Mu等,2018)。我们推测后者可能是观察到假阳性的来源。考虑到无论是正义还是反义RNA链都足以启动NASBA反应,且我们的NASBA引物在一个引物中包含T7 RNA聚合酶启动子,另一个引物中包含适体编码序列,少量的引物二聚体可能导致假阳性。在每种情况下,假阳性都小于预期的扩增子并用DFHBI(是一种 RNA适配体荧光探针,是绿色荧光蛋白 (GFP) 荧光团的模拟物,用于对活细胞中的 RNA 进行成像)染色,与引物来源的RNA产物相符(图2B,E;补充图S1)。
这个问题描述了一个实验过程,即NASBA(核酸扩增技术)的灵敏度很高,容易出现假阳性,可以通过调节反应条件来抑制这种情况。Apta-NASBA是一种使用荧光适配体对特定基因进行扩增的方法,这里使用了肠聚集性大肠杆菌的aggR基因作为例子。典型的NASBA反应在约2小时内产生荧光,并经常出现假阳性。通过加入Duplex40,这些假阳性得以抑制。
实验中人们尝试使用一种称为Duplex40的40bp双链DNA和另一种称为Duplex100的100bp双链DNA来抑制不希望的非特异性扩增现象。实验结果显示,Duplex40在10µM的浓度下可以完全抑制非模板介导的扩增,同时相对于没有Duplex40的反应,最终荧光强度也仅有微小的降低。而Duplex100在更低的浓度(0.316µM)下也能有效地抑制假阳性,且效果相似。这表明,Duplex40和Duplex100都可以作为抑制剂,有效地控制某些扩增反应中的非特异性扩增现象。
实验中探索了所需的竞争双链DNA的浓度(以碱基对计),以抑制假阳性。随着长度的增加,所需浓度降低,Duplex40的最小浓度为400µM bp,而Duplex100的最小浓度为31.7µM bp。作者因此推测,T7 RNA聚合酶可能与核酸的结合时间与其长度成正比,并且非常长的核酸可以作为假阳性最有效的抑制剂。然而,实际情况并非如此,基因组(牛胸腺)DNA并没有抑制假阳性(补充图S4)。此外,作者还筛选了肝素,另一种高度负电荷的多电解质和T7 RNA聚合酶抑制剂,以较大水平使用Spinach标记的Apta-NASBA结构进行实验(补充图S5;Sastry和Ross, 1997; Capila和Linhardt, 2002; Paige等,2011)。肝素使正信号和假阳性信号的最大信号都降低了大约相同的程度。
我们还筛选了先前报道的Mg2+浓度,以抑制未受驱动的转录(Mu等人,2018)。不含Duplex40和Mg2+从12mM减少到10mM的反应也减少了假阳性信号,尽管阳性反应中的信号减少了(图2D)。进一步降低到8mM Mg2+完全抑制了无动力的转录,最终荧光强度进一步降低(图2D),导致阳性反应只能通过PAGE上的弱条带检测到(图2E)。T7 RNA聚合酶在启动转录时与在延伸过程中表现出不同的确认。如先前推测的那样,高Mg2+浓度促进T7 RNA聚合酶的延伸相关构象,允许在Apta NASBA反应中的引物上发生无动力转录(Mu等人,2018)。
我们使用Duplex40作为假阳性抑制剂对反应进行了进一步的优化。优化包括去除氯化物(补充图S6;Chamberlin和Ring 1973),改变引物浓度(补充图S7),添加渗透液(补充图S8-S13),改变钾浓度(补充图S14),以及添加无机焦磷酸酶(补充图S15;Cunningham和Ofengand 1990)。优化的反应条件导致1pM的检测极限(图2F),没有来自阴性反应(即缺乏模板的反应)的信号。低于1pM的浓度未能扩增。
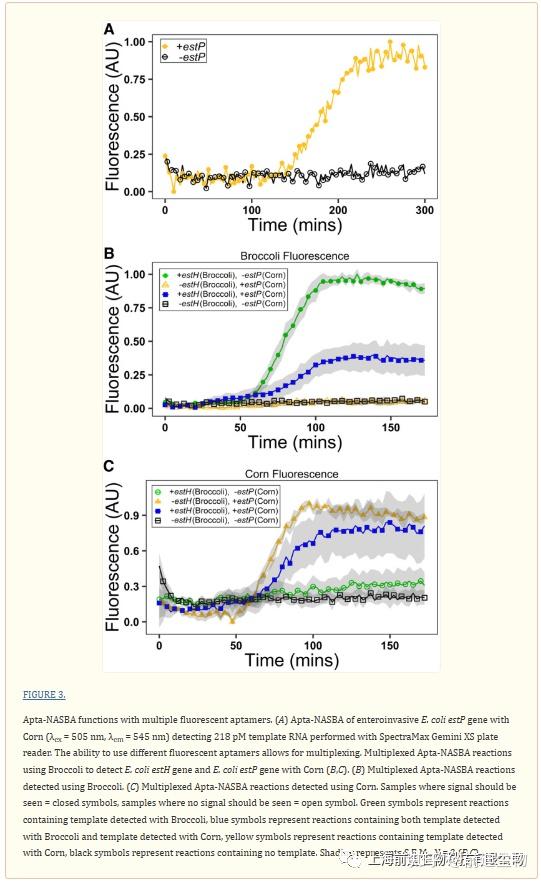
在观察到Apta NASBA系统与绿色适配体Broccoli(λex=472 nm,λem=507 nm)一起发挥作用后,我们试图使用黄色适配体Corn(λex=505 nm,λem=545 nm)将Apta NASBA扩展成不同的颜色(Song等人2017;Warner等人2017)。我们通过修改先前为实时PCR开发的引物,为另外两个病原体相关基因estP(使用玉米观察)和estH(使用西兰花观察)设计了引物。estP和estH产生在产肠毒素大肠杆菌中发现的两种不同的热稳定肠毒素变体,产肠毒素的大肠杆菌是旅行者和婴儿腹泻的主要原因(Grate和Wilson 1999;Plourde 2003;Hidaka等人2009)。此外,我们为aggR设计了另一种引物,使用孔雀绿适配体,一种红色适配体(λex=630nm,λem=650nm)进行观察(Grate和Wilson 1999;Babendure等人2003;Hidaka等人2009)。使用玉米检测estP和孔雀石绿适体检测aggR的反应是成功的(图3A;补充图S16)。与我们发现Duplex40提供的高度特异性一致,扩增子只有在用其同源引物组扩增时才表现出信号(补充图S17)。
考虑到严格的引物-扩增子关系,我们试图在多重检测系统中使用这些正交引物对。在这里,我们分别使用玉米和西兰花在单个反应中检测estP和estH(图3B,C)。我们还证明了使用西兰花、玉米和孔雀石绿适体的组合进行读出的双检测estP和aggR(补充图S18、S19)。
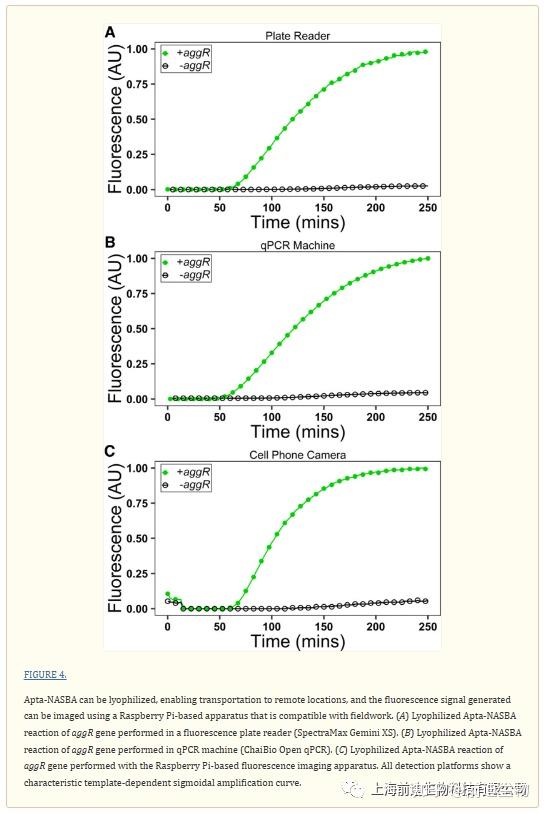
Apta NASBA的等温性质消除了对电源要求严格的热循环器的需求。为了进一步提高Apta NASBA在现场诊断和低资源领域的实用性,我们使用手机摄像头模块将冻干的Apta NASBA-反应与低成本的基于树莓派的检测平台(Hoog等人,2019)相结合(图4C)。编写了一个简短的Python脚本来控制荧光反应的图像采集,并根据像素值确定荧光强度(Hoog等人,2019)。在更昂贵的平板阅读器和基于qPCR的荧光检测平台上从Apta NASBA获得的荧光曲线在功能上与在基于Raspberry Pi的检测平台上观察到的荧光曲线相同(图4A–C)。我们发现,在添加冷冻保护剂(海藻糖)的情况下,Apta NASBA反应在冷冻干燥后是可行的,允许运输到没有冷链的偏远地区(图4C;补充图S20;补充视频S1)。
考虑到大量已知的荧光RNA折叠成G-Quadruplex结构,含有荧光标签的多结构域RNA的计算机辅助设计可能存在问题。我们观察到,尽管预测了错误的折叠,但折叠预测软件的使用通常能够成功预测给定RNA的结构域是否会独立折叠或诱导错误折叠。
本文中开发的RNA构建体为设计具有不同折叠的多结构域RNA提供了概念证明,包括二级结构预测算法不适应的折叠,特别是G-四重链。这些似乎在体外选择的荧光适体中普遍存在。例如,折叠预测算法显示Spinach是第一个发表的含有G-Quadruplex的荧光RNA适体,是一种茎环结构(Paige等人,2011)。然而,Spinach是现有折叠预测算法无法捕捉到的G-Quadruplex(Huang等人,2014;Warner等人,2014)。Apta NASBA读数中G-Quadruplex的存在是钾增加导致更大信号的原因。单价阳离子是正确的G-Quadruplex折叠和稳定性所必需的。在其他几种荧光适体中也观察到了多样和新颖的折叠。本文的工作为任意RNA的设计和部署提供了重要的概念证明,包括那些形成结构预测软件未考虑的非经典二级结构的RNA。
Apta NASBA代表了一个有吸引力的病原体检测平台,能够在偏远、低资源地区进行检测。鉴于其模块性和与一管多色序列检测的兼容性,它有望成为一个高度可扩展的平台。在目前的工作中,使用的读数是荧光适体。考虑到功能性RNA是使用第二引物安装的,Apta NASBA平台是高度通用的,并且存在安装任意核酸的可能性,包括其他功能性RNA,例如用于调节无细胞蛋白质表达系统的适体,或小转录激活RNA(STARs)(Chappell等人,2015)。
四、材料和方法
寡核苷酸从Integrated DNA Technologies获得,脱盐后按原样使用。所使用的寡核苷酸序列在补充表S1中给出。酶来自新英格兰生物实验室(Ipswitch),但T7 RNAP和mMuLV除外,它们在内部过表达,如下所述。
NASBA反应
NASBA是在对Sookanan、van Gemen和Malek最初报道的程序进行广泛修改的基础上进行的(Sookananan等人,1995)。反应的总体积为25µL。制备5×缓冲液,其包含200mM乙酸三酯、420mM KOAc、60mM MgOAc、dATP、dTTP、dGTP和dCTP各5mM、ATP、UTP、GTP和CTP各10mM。制备含有T7 RNAP、Moloney小鼠白血病病毒逆转录酶(mMuLV)、RNase H(New England Biolabs股份有限公司,M0297)和牛血清白蛋白(New EnglandBiolabs股份有限公司,B9000S)的酶混合物。存在的酶的最终反应浓度/量为每25µL反应1µM T7 RNAP、0.25µM mMuLV、0.2 U RNase H和2.6µg牛血清白蛋白。将5×缓冲液添加到含有1µM引物1、1µM底漆2、100µM同源染料、1 mM DTT、166 mM KOAc、10µM Duplex40、400 mM海藻糖、50µg无机焦磷酸酶(Bayou Biolabs)(Heili等人,2018)和4 mM MgOAc的溶液中,最终浓度为1×。不含酶混合物的反应在65°C下加热5分钟,然后在40°C加热5分钟。然后将酶混合物添加到反应中,并在40°C下孵育反应。所有适体都在低于其熔融温度的温度下使用。对于冻干反应,将除扩增子和染料外的全部反应在液氮中快速冷冻,并使用LABCONCO FreeZone 1升台式冷冻干燥系统冷冻干燥过夜,然后再悬浮在扩增子和染色液中。未对冻干反应进行65°C步骤。未添加模板的反应用作阴性对照。通过从每次测量中减去最小值,然后除以最大值,对仪器响应进行归一化,从而在每张图中对荧光光谱进行归一化。使用的模板在体外转录如下所述。引物序列见补充表S1。引物改编自Hidaka等人(2009)先前发表的引物。Geneious 10.1.3用于辅助引物设计。
T7 RNAP和mMuLV RT的过表达和纯化
用含有pT7-911Q(T7 RNAP)(Ichetovkin等人,1997)或pET MRT(mMuLV RT)(Yasukawa等人,2010)的大肠杆菌DH5α接种含有100µg/µL羧苄青霉素的10 mL LB。培养物在37°C下生长过夜,然后用于接种额外的1L含有100µg/µL羧苄青霉素的LB,并在37℃下生长至OD600在0.5和1之间。然后用1mM IPTG诱导培养物,并在37°C下生长3小时。将培养物在冰上冷却20分钟,并在3700 RPM下制粒15分钟。将颗粒在液氮中快速冷冻,并在−80°C下冷冻过夜。将颗粒在冷藏室中保持30分钟,然后溶解在20mL裂解缓冲液(50mM HEPES-KOH pH 7.6,1M NH4Cl,10mM MgCl2,7mM BME)中。将颗粒在裂解缓冲液中孵育30分钟,然后进行尖端超声处理。以50%的功率以15秒的间隔进行超声处理,直到施加了2kJ的总能量,然后使样品冷却5分钟。这一共重复了四次。然后将颗粒在4°C下以15000g离心45分钟。
将上清液应用于0.6mL Ni-NTA琼脂糖珠(GoldBio,H-350-50)上,并在冷藏室的摇床上孵育1小时。以分批法进行洗涤和洗脱步骤。珠粒用10mL洗涤缓冲液洗涤10分钟,然后再次用10mL冲洗缓冲液(50mM HEPES pH 7.6、1M NH4Cl、10mM MgCl2、15mM咪唑、7mM BME)洗涤15分钟。将一定量的3mL洗脱缓冲液(50 mM HEPES-KOH pH 7.6、100 mM KCl、10 mM MgCl1、300 mM咪唑、7 mM BME。使用Slide-A-Lyzer Dialysis Cassette,2000 MWCO(Thermo Fisher Scientific,66203),在500 mL 2×储存缓冲液(100 mM Tris-HCl pH 7.6,200 mM KCl,20 mM MgCl2,14 mM BME)中透析洗脱过夜,然后在额外的500 mL 2倍储存缓冲液中透析3小时。在不含甘油的相同储存缓冲液中将用于冷冻干燥的酶制备。
使用NanoDrop ND-1000使用计算的A280对蛋白质进行定量。蛋白质活性通过西兰花适体的体外转录和在荧光平板读取器(T7 RNAP)上的动力学监测或estH基因的逆转录和在SYBR金染色的尿素PAGE凝胶(mMuLV RT)上的可视化来评估。
模板转录
进行一个millileter转录以获得用于NASBA反应的模板。反应物含有40mM乙酸三酯、24mM谷氨酸镁、100mM谷氨酸钾、2mM亚精胺、1mM DTT、4mM ATP、4mM GTP、4mM UTP、4mM CTP、1µM过表达的T7 RNA聚合酶(相反,可以使用新英格兰生物实验室T7 RNA聚酶M0251L,并且根据补充表S1使用双链DNA模板(Heili等人,2018)。最佳模板浓度是根据经验确定的,这里使用的扩增子通常为~100nM。反应物在37°C下孵育8小时。对于转录物纯化,首先用1 U/µg Turbo DNase(Thermo Fisher Scientific,S11494)在37°C下处理反应15分钟,然后用乙醇沉淀。将沉淀的RNA重悬于1×负载缓冲液(1×TBE,8M尿素)中,并在10%尿素PAGE凝胶上分离;通过荧光TLC板上的UV阴影切除全长产物带。通过在1至3 mL 1×TBE中迫使凝胶切片通过21 Ga针,然后在液氮和55°C水浴之间交替进行3次冻融循环,对凝胶切片进行物理干扰。将含有RNA的TBE倾析,并进行乙醇沉淀。将最终产物悬浮在不含RNase的水中,并使用NanoDrop ND-1000通过A260测定浓度。
小牛胸腺DNA
小牛胸腺DNA(Sigma D1501-100MG)在10mM Tris乙酸酯(pH 8)、1mM EDTA中稀释至2mg/mL,并旋转直至均匀。为了最大限度地减少剪切,没有进行涡流。为了测定所用DNA的量,使用NanoDrop ND-1000通过A260测定溶液浓度为2.32mM。
凝胶染色
将10%的尿素PAGE凝胶(1×TBE,8M尿素)在两种不同的折叠缓冲液(100mM乙酸三酯pH 8,10mM MgCl2,500mM KCl)中孵育20分钟,每次从凝胶中去除尿素,并提供K+和Mg2+以支持适体折叠。然后将凝胶在掺有10µM DFHBI的折叠缓冲液中孵育15分钟。如果凝胶上的荧光背景很高,则将其在折叠缓冲液中将其去除5分钟。使用橙色滤光片将凝胶成像在凝胶doc(Omega Lum G,Aplegen)上。为了显示不含适体的序列,将凝胶在1×SYBR Gold中再染色15分钟,并在相同条件下重新老化。
微信扫一扫
关注该公众号
回复
支持
反对
举报
长长的路
长长的路
当前离线
金桔
金币
威望
贡献
回帖
0
精华
在线时间
小时
发表于 2024-9-21 10:28
|
显示全部楼层
LAMP 是2000年日本研究人员Notomi等发明的
回复
支持
反对
举报
队长是我
队长是我
当前离线
金桔
金币
威望
贡献
回帖
0
精华
在线时间
小时
发表于 2024-9-21 10:28
|
显示全部楼层
诶……恒温pcr(isothermal pcr)技术有很多种,很多很多种,LAMP,桥式pcr,EXPAR,RCA……
而且我记得LAMP是日本人发明的,石磊顶多算是做了个应用,讲道理做技术原理的略微看不上这种拿别人的技术用了一下就说是自己独创的……
回复
支持
反对
举报
返回列表
发表回复
高级模式
B
Color
Image
Link
Quote
Code
Smilies
您需要登录后才可以回帖
登录
|
立即注册
本版积分规则
发表回复
回帖后跳转到最后一页
关闭
官方推荐
/3
AI助手<小桔子>来了!
欢迎来交流,可以回答IVD行业各类问题!
查看 »
IVD业界薪资调查(月薪/税前)
长期活动,投票后可见结果!看看咱们这个行业个人的前景如何。请热爱行业的桔友们积极参与!
查看 »
小桔灯网视频号开通了!
扫描二维码,关注视频号!
查看 »
返回顶部
快速回复
返回列表
客服中心
搜索
洽谈合作
关注微信
微信扫一扫关注本站公众号
个人中心
个人中心
登录或注册
业务合作
-
投稿通道
-
友链申请
-
手机版
-
联系我们
-
免责声明
-
返回首页
Copyright © 2008-2024
小桔灯网
(https://www.iivd.net) 版权所有 All Rights Reserved.
免责声明: 本网不承担任何由内容提供商提供的信息所引起的争议和法律责任。
Powered by
Discuz!
X5.0 技术支持:
宇翼科技
浙ICP备18026348号-2
浙公网安备33010802005999号
快速回复
返回顶部
返回列表

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 






 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 发表于 2024-9-21 10:26
发表于 2024-9-21 10:26


 发表于 2024-9-21 10:27
发表于 2024-9-21 10:27