用户名
UID
Email
密码
记住
立即注册
找回密码
只需一步,快速开始
微信扫一扫,快速登录
开启辅助访问
快捷导航
门户
Portal
社区
BBS
资讯
会议
市场
产品
问答
数据
专题
帮助
签到
每日签到
企业联盟
人才基地
独立实验室
产业园区
投资机构
检验科
招标动态
供给发布
同行交流
悬赏任务
共享资源
VIP资源
百科词条
互动话题
导读
动态
广播
淘贴
法规政策
市场营销
创业投资
会议信息
企业新闻
新品介绍
体系交流
注册交流
临床交流
同行交流
技术杂谈
检验杂谈
今日桔说
共享资源
VIP专区
企业联盟
投资机构
产业园区
业务合作
投稿通道
升级会员
联系我们
搜索
搜索
本版
文章
帖子
用户
小桔灯网
»
社区
›
C、IVD技术区
›
基因编辑技术
›
有无大神能够从进化的角度推测或者谈谈张峰今年发现的真 ...
图文播报
2026庆【网站十三周
2025庆【网站十二周
2024庆中秋、迎国庆
2024庆【网站十一周
2023庆【网站十周年
2022庆【网站九周年
返回列表
查看:
9551
|
回复:
3
[分享]
有无大神能够从进化的角度推测或者谈谈张峰今年发现的真核生物中的fanzor蛋白到底有什么意义?
[复制链接]
非诚勿扰孟非
非诚勿扰孟非
当前离线
金桔
金币
威望
贡献
回帖
0
精华
在线时间
小时
发表于 2024-9-11 20:11
|
显示全部楼层
|
阅读模式
登陆有奖并可浏览互动!
您需要
登录
才可以下载或查看,没有账号?
立即注册
×
真核生物中的fanzor和wRNA,类crispr cas9的基因编辑系统?
原文地址:https://www.zhihu.com/question/626229415
回复
举报
卡卡
卡卡
当前离线
金桔
金币
威望
贡献
回帖
0
精华
在线时间
小时
发表于 2024-9-11 20:11
|
显示全部楼层
如题,最近看到一篇2013年的文献:“
Homologues of bacterial TnpB_IS605 are widespread in diverse eukaryotic transposable elements
”
文献中,作者发现了两组 TnpB 样蛋白(Fanzor1 和 Fanzor2),它们广泛存在于多种
真核转座元件(TE)
和感染真核生物的大型双链 DNA(dsDNA)病毒中。TnpB/Fanzor 蛋白的系统发育和分布表明
它们可能通过病毒在真核物种中传播。这篇文章其实非常的重要,直接启发了张峰教授在2023年发表的利用alphafold2来搜索真核生物体内可用的基因编辑新系统的文章。
但是此前,本人对转座子实在是毫无记忆,毕竟大学已经过去了快10年了,所以一份小小的前言,就阅读了好久。
转座子(TE) 的分类
可转座元件(Transposable Elements,TEs)主要分为两大类:
DNA转座元件
和
RNA转座元件(逆转录转座子)
。
1.
DNA转座元件
:这类转座元件直接通过DNA中介物进行转座,不需要RNA中介。它们可以进一步分为两个亚类:复制/粘贴转座元件和剪切/粘贴转座元件。复制/粘贴转座元件,如转座子,通过复制自身并将副本插入基因组的新位置。剪切/粘贴转座元件,如IS元件和Tn元件,通过从原始位置剪切并粘贴到新位置。
2.
类RNA转座元件
:这类转座元件通过RNA中介物进行转座,需要逆转录酶将RNA转录为DNA,然后插入基因组。它们主要包括
长终端重复(LTR)逆转录转座子
,如逆转录转座子,和
非LTR逆转录转座子
,如长散在重复序列(LINEs)和短散在重复序列(SINEs)。
这些转座元件在基因组中占据了大量的空间,对基因组的结构和功能有重要影响。在生物信息学研究中,对转座元件的研究有助于理解基因组的演化和多样性。
逆转录酶和转座酶
通常,转座元件仅编码其
复制和插入
所必需的蛋白质,包括
逆转录酶
和
转座酶
(Tpases)
1. 逆转录酶:逆转录酶是一种酶,它可以将RNA模板转录为DNA,这是一个与常规的DNA到RNA的转录过程相反的过程,因此得名“逆转录”。逆转录酶在类RNA转座元件的复制和插入过程中起着关键作用。这些转座元件首先被转录为RNA,然后由逆转录酶将RNA转录为DNA,最后将这些DNA插入到基因组的新位置。
2. 转座酶(Tpases):转座酶是一种可以切割DNA并将可转座元件插入新位置的酶。在类DNA转座元件的复制和插入过程中,转座酶起着关键作用。例如,在
剪切/粘贴
类型的转座元件中,转座酶会将转座元件从其原始位置切割出来,然后将其粘贴到基因组的新位置。
转座酶的分类
可转座元件(TEs)编码的转座酶主要有四种类型:
1. DDE/D:这是最常见的转座酶类型,以其活性位点上的三个关键氨基酸——天冬氨酸(D)、天冬氨酸(D)和谷氨酸(E)或天冬氨酸(D)命名。这些酶由大多数细菌插入序列 (IS)、真核 DNA 转座子和 LTR 逆转录转座子编码。
2. RT:以逆转录酶 (RT) 为代表,由多种非 LTR 和 LTR 逆转录转座子编码。
3. YR(Tyrosine Recombinases):这种类型的转座酶以其活性位点上的两个关键酪氨酸(Y)残基命名。这些酶主要由 IS91、Helitron、IS200/IS605、Crypton 和 DIRS-逆转录转座子家族编码。
4. SR(Serine Recombinase):这种类型的转座酶以其活性位点上的关键氨基酸——丝氨酸(S)命名。这些酶主要由 IS607 家族、Tn4451 和噬菌体 phiC31 编码。
这四种类型的转座酶都能催化TEs从一个基因组位置移动到另一个位置的过程,但它们的工作机制和特性各不相同。在生物信息学研究中,对这些转座酶的研究有助于理解TEs如何影响基因组的结构和功能。
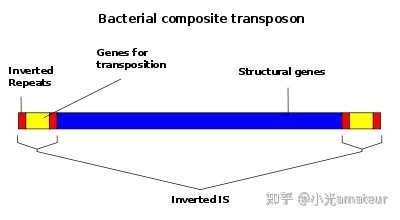
DNA转座子的结构和转座机制
大多数 DNA 转座子两侧都有
末端反向重复序列
(TIR) 和
靶位点重复
(TSD),并通过 DDE 转座酶使用的“剪切和粘贴”机制进行转座,尽管有些转座子使用复制机制 (Tn3) ,或者能够切换到复制模式(例如,MuDr、Tn7 和 IS903)。其转座的过程通常涉及以下几个步骤:
1. 识别
:转座子的转座酶(例如转座酶TnpA)识别转座子的特定DNA序列,这些序列通常是转座子
两端的末端反向重复序列(TIRs)
。
2. 切割
:转座酶切割转座子两端的DNA,从而释放转座子。在剪切/粘贴转座子中,这一步涉及到从原始位置切割整个转座子;而在复制/粘贴转座子中,转座子被复制,原始转座子保留在原位。
3. 转移
:释放的转座子或其复制品随后被转移至新的位置。在这个过程中,转座子可能通过转座酶与宿主因子的相互作用来定位到目标位点。
4. 插入
:转座子插入新位置时,通常会在插入点两侧产生切口,这些切口随后被宿主的DNA修复机制修复。在修复过程中,
会在新位置两侧产生靶位点重复序列
(TSDs)。
5. 修复
:最后,宿主的DNA修复系统修复插入点两侧的断裂,完成转座子的插入过程。
但是要注意的是,使用 YR 和 SR 作为 Tpase 的转座子缺乏 TIRs,并且在插入时不产生 TSDs。然而,它们的末端发夹结构(IS200/605 家族)或末端短同向重复序列(Crypton)对于转座很重要。
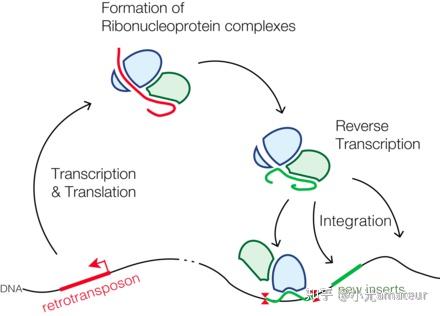
逆转录转座子的结构和转座机制
逆转录转座子是一类通过RNA中介物进行转座的可转座元件(TEs),它们的结构和转座机制具有以下特点:
结构
1
. 长终端重复(LTR)
:许多逆转录转座子的两端具有长终端重复序列,这些序列在转座过程中起到关键作用。
2. 逆转录酶(Reverse Transcriptase, RT)
:逆转录转座子编码逆转录酶,这是它们转座机制中的核心酶。
3. 整合酶(Integrase, IN)
:某些逆转录转座子还编码整合酶,它有助于将逆转录后的DNA整合到宿主基因组中。
4. RNA结合蛋白
:逆转录转座子可能编码与RNA结合的蛋白质,这些蛋白质参与转座子RNA的稳定和转座。
5. 其他蛋白质
:一些逆转录转座子还可能编码其他功能蛋白,如蛋白酶(Protease)和核糖体结合位点(Ribosome Binding Site, RBS)等。
转座机制
1. 转录
:逆转录转座子首先被宿主细胞的转录机制转录为RNA。
2. 逆转录
:逆转录酶将转录出的RNA逆转录为单链DNA。
3. 二级DNA合成
:逆转录后的单链DNA被转化为双链DNA,这一步骤通常涉及宿主细胞的DNA修复和复制机制。
4. 整合
:整合酶将双链DNA整合到宿主基因组的新位置,这个过程可能产生靶位点重复序列(TSDs)。
5. 表达调控
:逆转录转座子的表达可能受到宿主基因组中的调控元件的影响,如启动子和增强子。
TnpA, TnpB和Fanzor
TnpA是一种转座酶
,它在某些类型的可转座元件(TEs),特别是IS200/IS605家族的转座子中发现。TnpA通常负责催化转座子从一个基因组位置切割并插入到另一个位置的过程。这种酶识别特定的DNA序列,通常是转座子的末端反向重复序列(TIRs),然后通过切割DNA并介导转座子的移动来执行其功能。
在IS200/IS605家族的转座子中,TnpA与另一种
蛋白TnpB
共同作用,可能参与转座过程的不同阶段。TnpA的活性和特异性可能受到TnpB的调节,
尽管TnpB的确切作用尚不完全清楚
。TnpA通常具有特定的催化活性位点,这些位点对于其切割和插入DNA的能力至关重要。
针对 IS607、ISHp608 和 ISDra2 元件(后两者属于 IS200/IS605 家族)的三个独立实验表明,
TnpB 对于大肠杆菌 和耐辐射球菌 中的转座是可有可无的
。有趣的是,
许多 IS 元件(例如 IS1341、IS809 和 IS1136)将 TnpB 编码为唯一的蛋白质(假定的转座酶),但 TnpB 介导的转座的支持证据仍然缺失
。与 IS200/IS605 和 IS607 家族的其他元件一样,这些仅 TnpB 的转座子缺乏 TIR 和 TSD。
一种可能性是这些元件代表 IS607 或 IS200/IS605 样转座子的非自主衍生物,其中 TnpA 被删除。
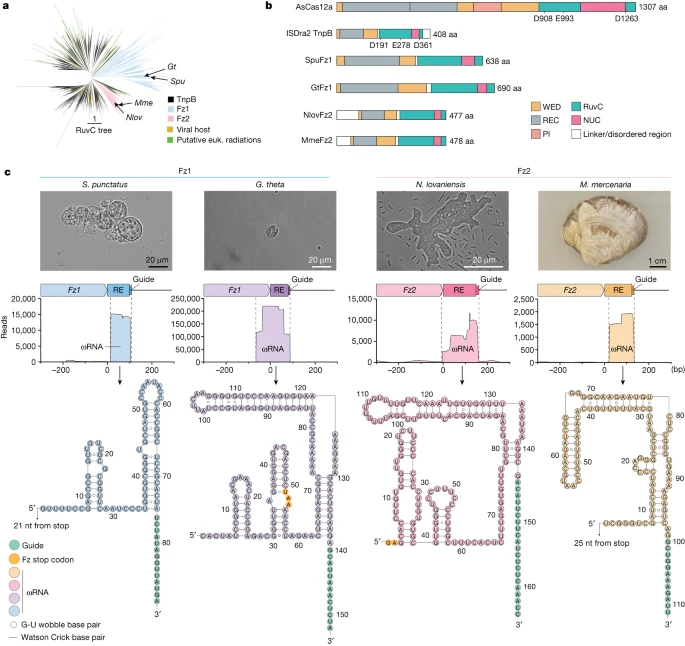
Fanzor包括(Fanzor1 和 Fanzor2)其实是TnpB类似蛋白质,
Fanzor 和 TnpB 蛋白
均包含一组从蛋白中心延伸至 C 末端的严格保守残基
,D-X(125, 275)-[TS]-[TS]-X-X-[C4 锌指]-X(5, 50)-RD。 C4 锌指在保守域数据库 (CDD) 中称为 OrfB_Zn_ribbon ([CDD:pfam07282])。从系统发育角度来看,
Fanzor1 蛋白形成一个单独的进化枝
,而
Fanzor2 蛋白与来自 IS607 家族的一小组细菌 TnpB 蛋白共簇
。 Fanzor1 蛋白被来自至少五个不同超家族的转座元件捕获:Mariner、Sola、IS4、Helitron 和 MuDr。 Fanzor2 蛋白由 IS607 型转座子编码。虽然目前尚不清楚 Fanzor/TnpB 蛋白的生物学功能,但有迹象表明 Fanzor1 蛋白可能具有甲基转移酶的功能。这是基于 PGv-1、Mariner-2 _PGv 和 Mariner-1 _OLpv 三个元件的比较,每个元件编码三种蛋白质,包括 Mariner-Tpase、核酸内切酶以及甲基转移酶或 Fanzor1 蛋白质。
病毒可能有助于 Fanzor 蛋白在真核生物中的传播。
回复
支持
反对
举报
长长的路
长长的路
当前离线
金桔
金币
威望
贡献
回帖
0
精华
在线时间
小时
发表于 2024-9-11 20:12
|
显示全部楼层
想知道关于转座子角度的猜测和联系甲基转移酶相关的猜想
回复
支持
反对
举报
清风寡欲
清风寡欲
当前离线
金桔
金币
威望
贡献
回帖
0
精华
在线时间
小时
发表于 2024-9-11 20:12
|
显示全部楼层
真核生物的 Fanzor 蛋白(Fz)由
转座元件
编码,系统发育分析支持 Fz1、Fz2 分别起源于不同的细菌 TnpB,大概是在病毒搬运下从细菌彼此独立地
水平基因转移
而来、在真核生物间由病毒和真核共生体继续转移,这些转移可能现在仍在发生[1]。
已测序的多种真核生物和一些感染真核生物的病毒(尤其是感染与细菌共生的变形虫的巨型病毒)现在含有类 TnpB 的序列,一些病毒现在含有 Fz 序列,体现了不同时期发生的不同方向的水平基因转移。
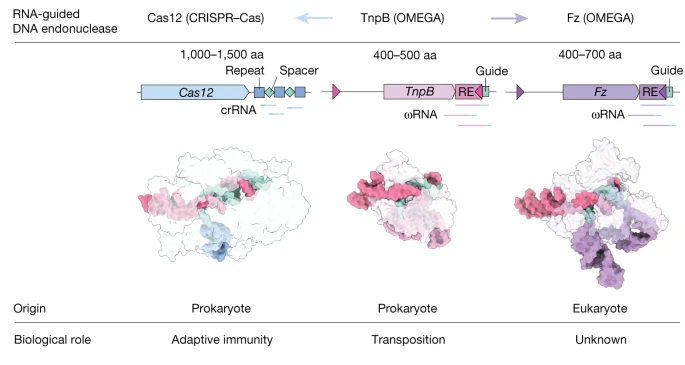
10.1038/s41586-023-06356-2
2013 年,一项研究认为 Fz 的结构和功能类似原核生物的 TnpB-IS200/IS605,二者可能通过甲基转移酶活性调节转座元件的活性;2021 年,两项研究认为原核生物的 TnpB、IscB、IsrB 蛋白是 obligate mobile element–guided activity(OMEGA)系统的核酸内切酶。OMEGA 系统可能是 CRISPR-Cas 系统的祖先,Cas12 可能起源于 TnpB。张峰团队由此推测 Fz 可能有核酸内切酶活性,进行实验。
实验显示,来自土壤真菌
Spizellomyces punctatus
等的几种 Fz 直系同源物在 omegaRNA 引导下剪切人基因组里 8 个位点的成功率不一,最高约 11.8%,低于 CRISPR-Cas 系统;测试了 111 种发生点突变的 Fz,其中几种剪切上述位点的成功率上升,最高约 18%;
Fz 在切割 DNA 靶点时不触发周围的核酸降解,这可能让其切割比一些 CRISPR 系统和 OMEGA 系统更精确、不易脱靶、不易造成附带影响。
Fz 的分子量较小,递送大概较容易。
Cas12a 蛋白比 Fz 和 TnpB 多几个结构域。omegaRNA 发挥与那些结构域类似的功能。
10.1038/s41586-023-06356-2
在可预见的未来,难以期待 Fz 系统的切割性能赶上 Cas9。Fz 可能发展为相对安全的基因编辑技术,
补充
CRISPR-Cas 技术。这未必能成功。
回复
支持
反对
举报
返回列表
发表回复
高级模式
B
Color
Image
Link
Quote
Code
Smilies
您需要登录后才可以回帖
登录
|
立即注册
本版积分规则
发表回复
回帖后跳转到最后一页
浏览过的版块
特定蛋白分析
关闭
官方推荐
/3
AI助手<小桔子>来了!
欢迎来交流,可以回答IVD行业各类问题!
查看 »
IVD业界薪资调查(月薪/税前)
长期活动,投票后可见结果!看看咱们这个行业个人的前景如何。请热爱行业的桔友们积极参与!
查看 »
小桔灯网视频号开通了!
扫描二维码,关注视频号!
查看 »
返回顶部
快速回复
返回列表
客服中心
搜索
洽谈合作
关注微信
微信扫一扫关注本站公众号
个人中心
个人中心
登录或注册
业务合作
-
投稿通道
-
友链申请
-
手机版
-
联系我们
-
免责声明
-
返回首页
Copyright © 2008-2024
小桔灯网
(https://www.iivd.net) 版权所有 All Rights Reserved.
免责声明: 本网不承担任何由内容提供商提供的信息所引起的争议和法律责任。
Powered by
Discuz!
X5.0 技术支持:
宇翼科技
浙ICP备18026348号-2
浙公网安备33010802005999号
快速回复
返回顶部
返回列表

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 






 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 发表于 2024-9-11 20:11
发表于 2024-9-11 20:11