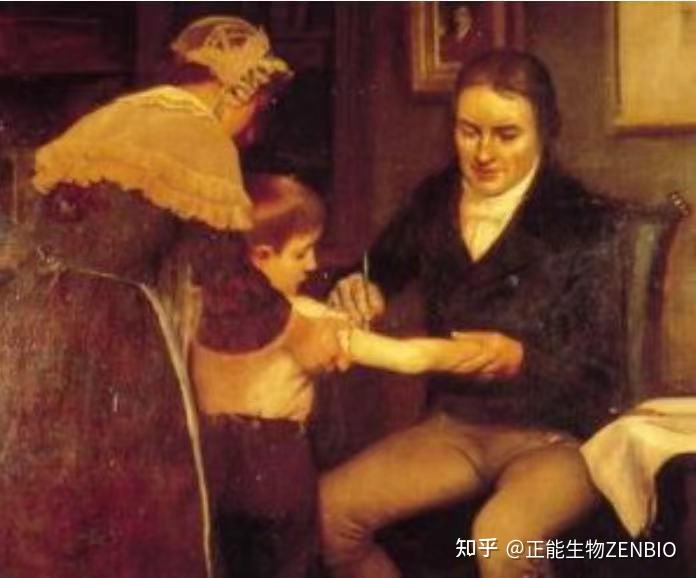
抗体这一概念,最早由Emil von Behring和Shibasabura Kitasato在1890年提出。他们在一篇具有里程碑意义的论文中,报道将白喉免疫过的动物血清注射到白喉感染的动物身上,让白喉感染的动物得到了治愈 [2],这一发现表明抗体在人类疾病防治方面具有巨大的潜力。Behring也因这项杰出的工作于1901年被授予诺贝尔奖。
Astrid Fagraeus在1948年描述了浆细胞特异性参与抗体的产生,随后Frank Burnet和David Talmage于1957年发展出了克隆选择学说。在Frank和David的理论中[4],淋巴细胞在遇到抗原之前会产生单一的特异性抗体分子,这与Linus Pauling在1940年提出的“诱导学说”相反,即抗原作为抗体的模板指导其合成[5]。

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 






 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 发表于 2024-9-8 22:54
发表于 2024-9-8 22:54