登陆有奖并可浏览互动!
您需要 登录 才可以下载或查看,没有账号?立即注册


×
自 2002 年被发现以来,炎症小体一直都是免疫和炎症疾病领域的热门选手,21 年来热度逐年攀升,时至今日,魅力丝毫不减~炎症小体究竟是何方神圣?为啥科研热度居高不下?

▐ 什么是炎症小体?“炎症小体”一词最早由 Jurg Tschopp 博士及其同事于 2002 年提出[1]。他们将炎症小体描述为一种“caspase 激活复合物”。炎症小体 (lnflammasomes) 是一种较大的多聚体蛋白复合物,主要存在于攻击病原体的先天免疫细胞中,它在感知外源性病原体或内源性危险信号时会启动炎症反应以及诱导细胞焦亡 。
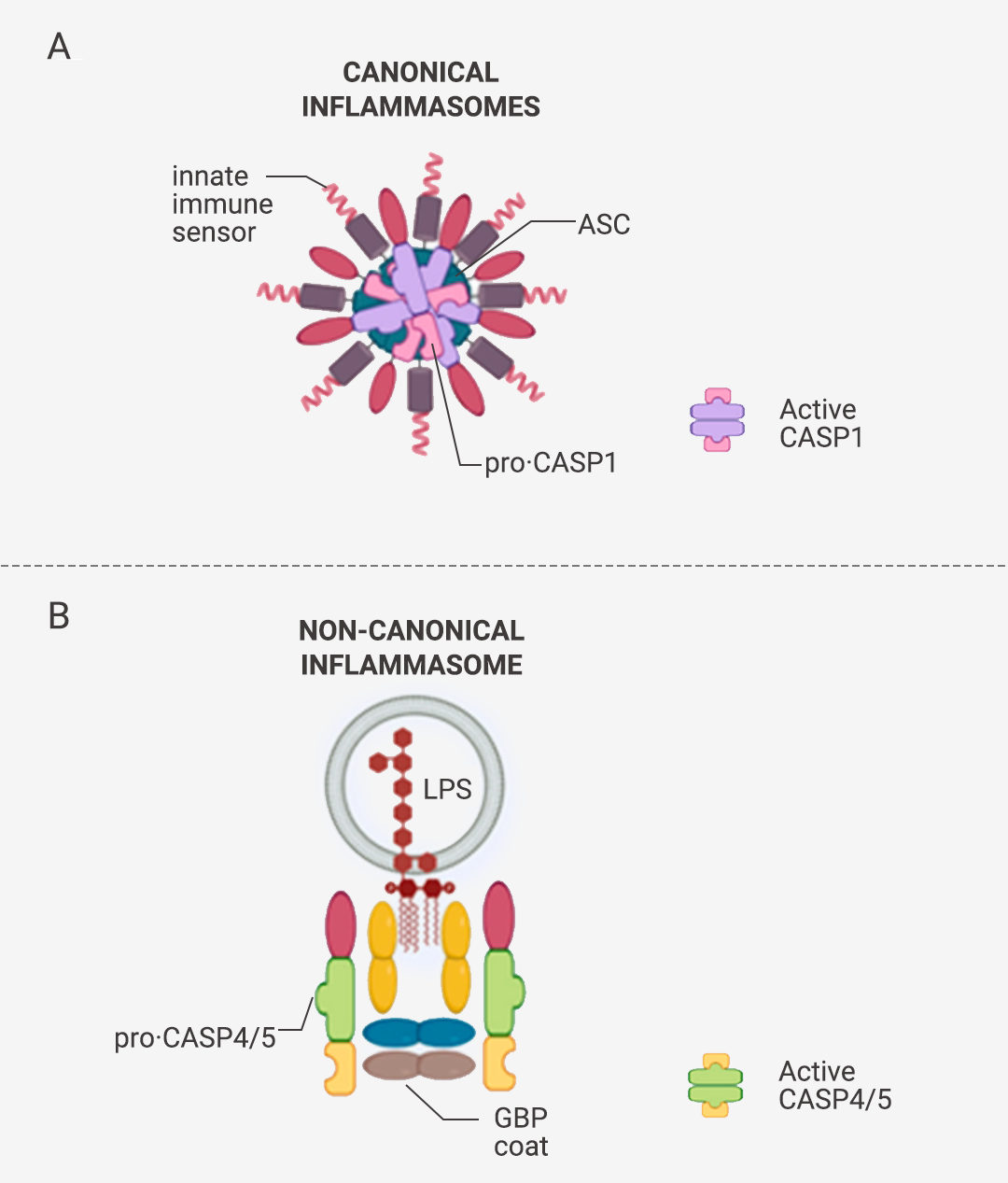
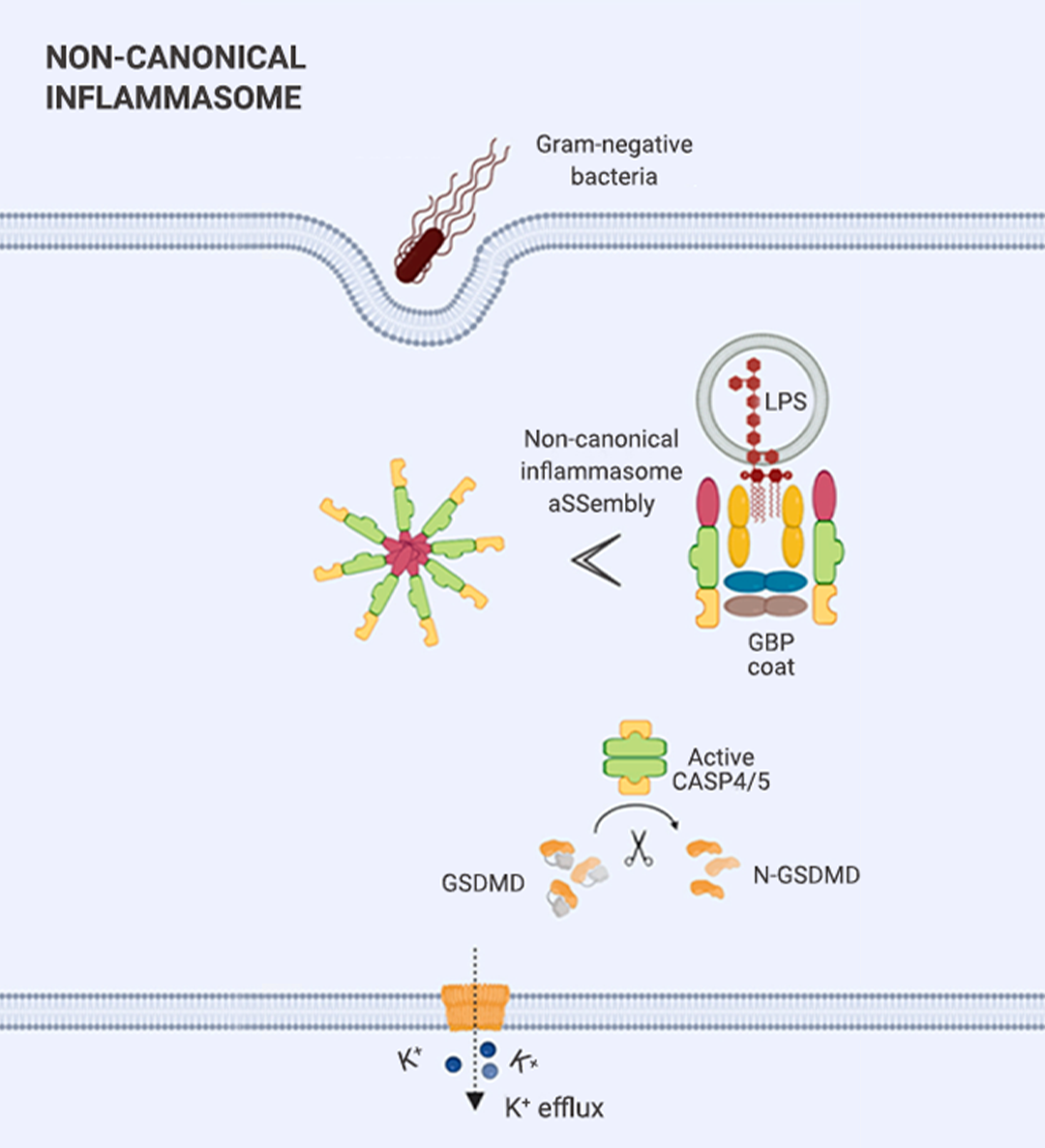
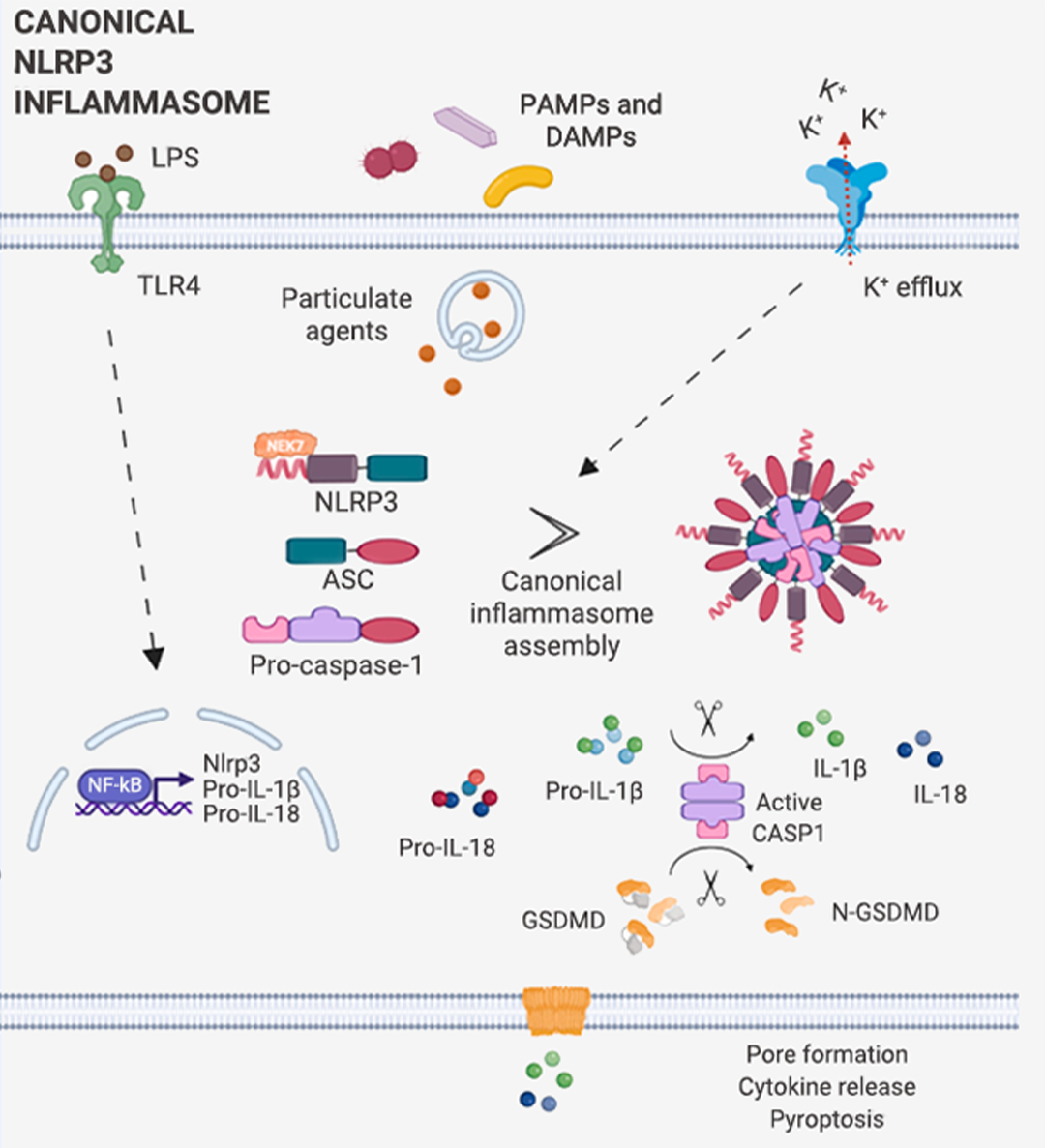
炎症小体分为两种:经典炎症小体和非经典炎症小体 (图 1)。
▐ 经典炎症小体
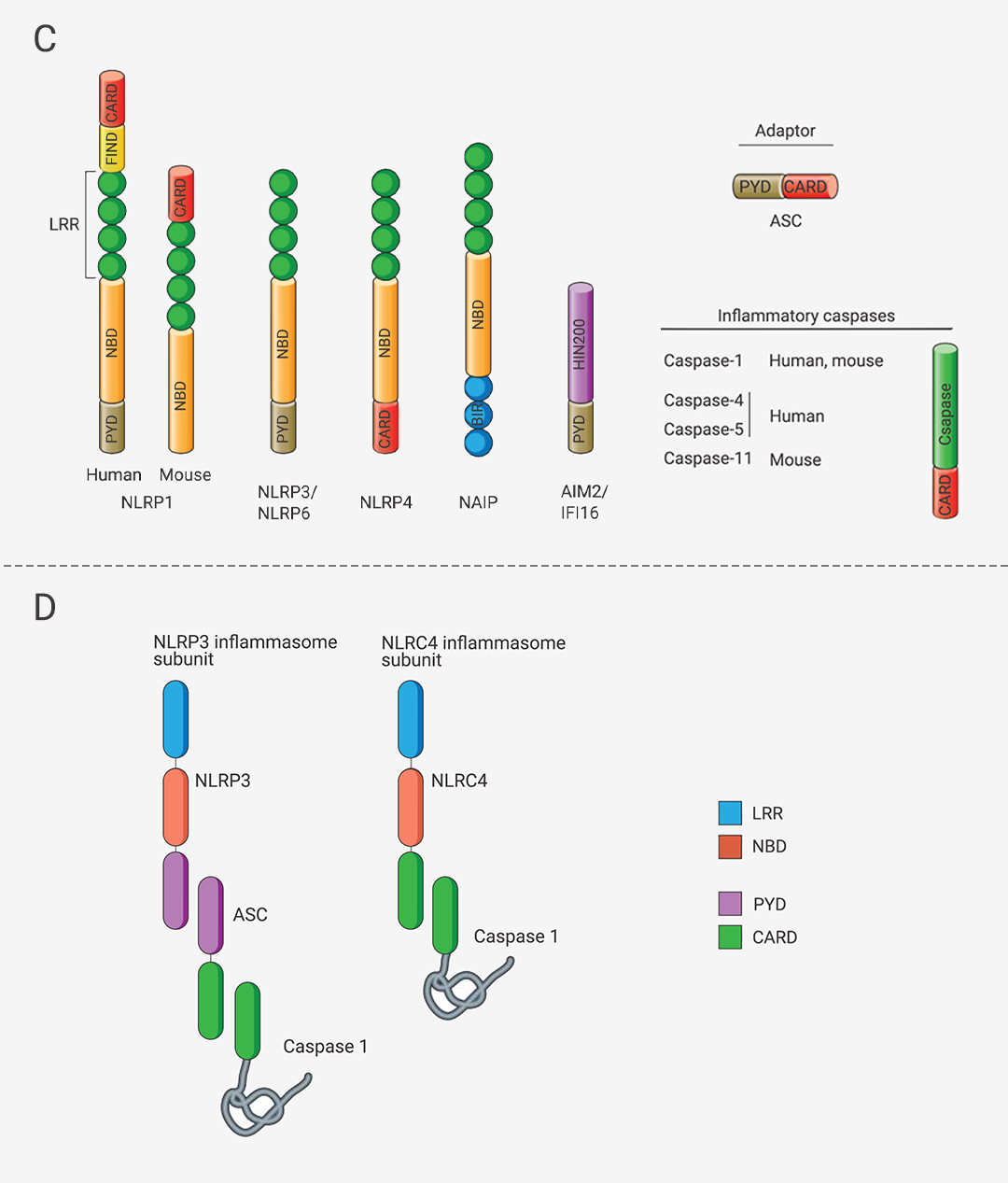
经典炎症小体是由一个感受器蛋白 (主要是 NLR 家族蛋白和 PYHIN 家族蛋白;常见上游感受器蛋白分类见表 1 [2][3][4])、一个衔接蛋白 ASC (NLRC4 炎症小体不需要 ASC) 和一个 Pro-Caspase-1 (Caspase 效应器) 通过 PYD (热蛋白结构域) 和/或 CARD (半胱天冬酶募集结构域) 结合在一起 (图 1C-D),形成的多聚体蛋白复合物 (图 1A)。图 1. 炎症小体组装、经典/非经典炎症体示意图[5][6][7]。 A. 经典炎症小体示意图;B. 非经典炎症小体示意图。C. 炎症小体组装部件包含的结构域。D. (以 NLRP3 炎症小体为例) ASC 通过 PYD 和 CARD 两个结构域,将感受器蛋白和 Caspase 效应器衔接在一起,组装成炎症小体;NLRC4 感受器蛋白有 CARD 结构域且无 PYD 结构域,故不需 ASC 即可直接与 Caspase 效应器结合。
▐ 非经典炎症小体
非经典炎症小体是由人类 Pro-Caspase-4/Pro-Caspase-5 (或它们的鼠直系同源 Pro-Caspase-11) 与 LPS 构成的大分子蛋白复合物 (图 1 B)。简单来说:
经典炎症小体:感受器蛋白 + ASC + Pro-Caspase-1。非经典炎症小体:Pro-Caspase-4/5/11 + LPS。
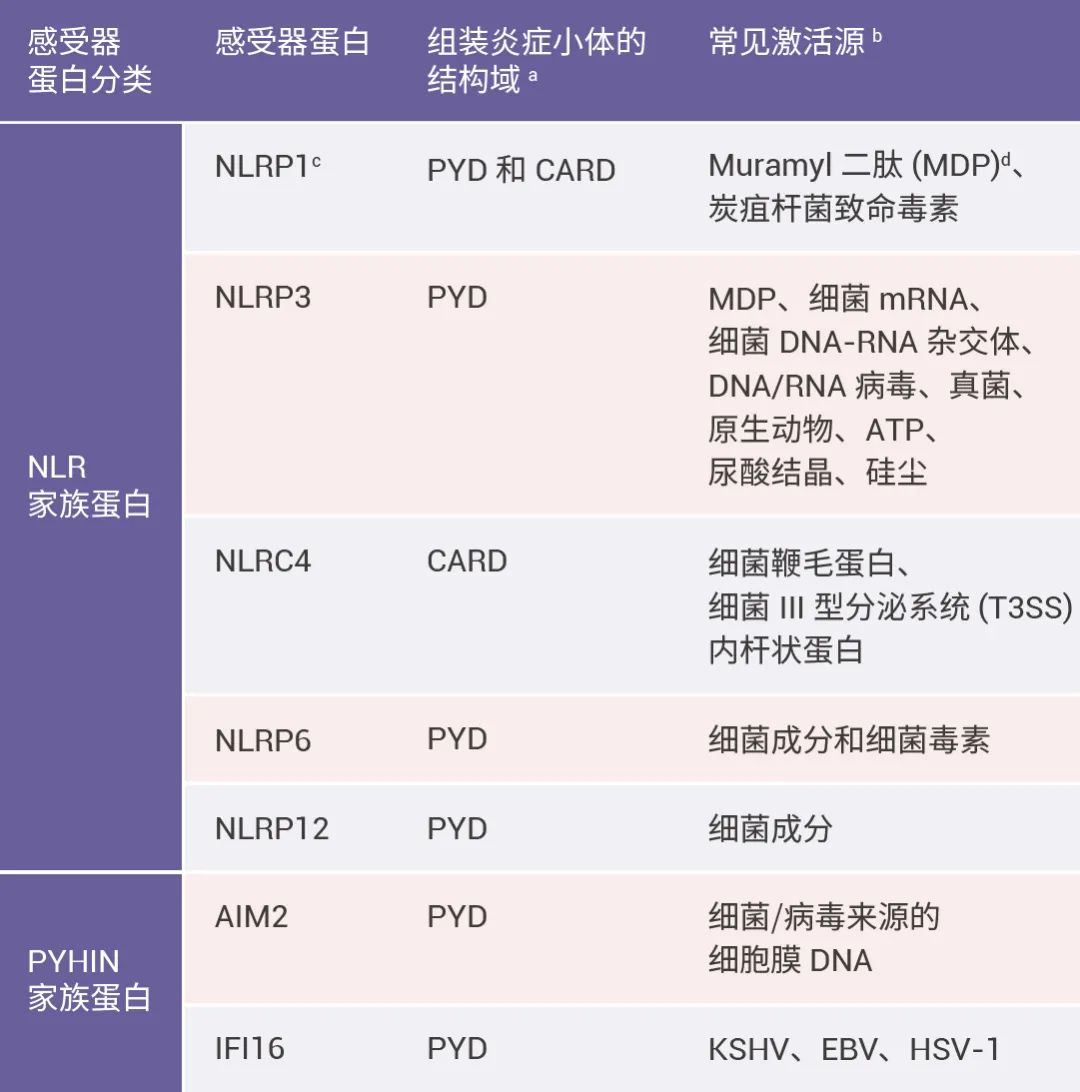
表 1. 经典炎症小体常见受器蛋白的分、结构域以及常见激活源[2][3][4]。 注:a. 由于炎症小体组装涉及的结构域为 PYD 和 CARD,故表中仅列出感受器蛋白中是否有这两种结构域;b. 感受器蛋白不一定需要直接与这些信号结合;c. NLRP1 是 被发现的炎症小体感受器蛋白;d. Muramyl 二肽 (MDP) 是一种来自革兰氏阳性和阴性细菌的肽聚糖片段。 被发现的炎症小体感受器蛋白;d. Muramyl 二肽 (MDP) 是一种来自革兰氏阳性和阴性细菌的肽聚糖片段。

▐ 经典炎症小体活化通路
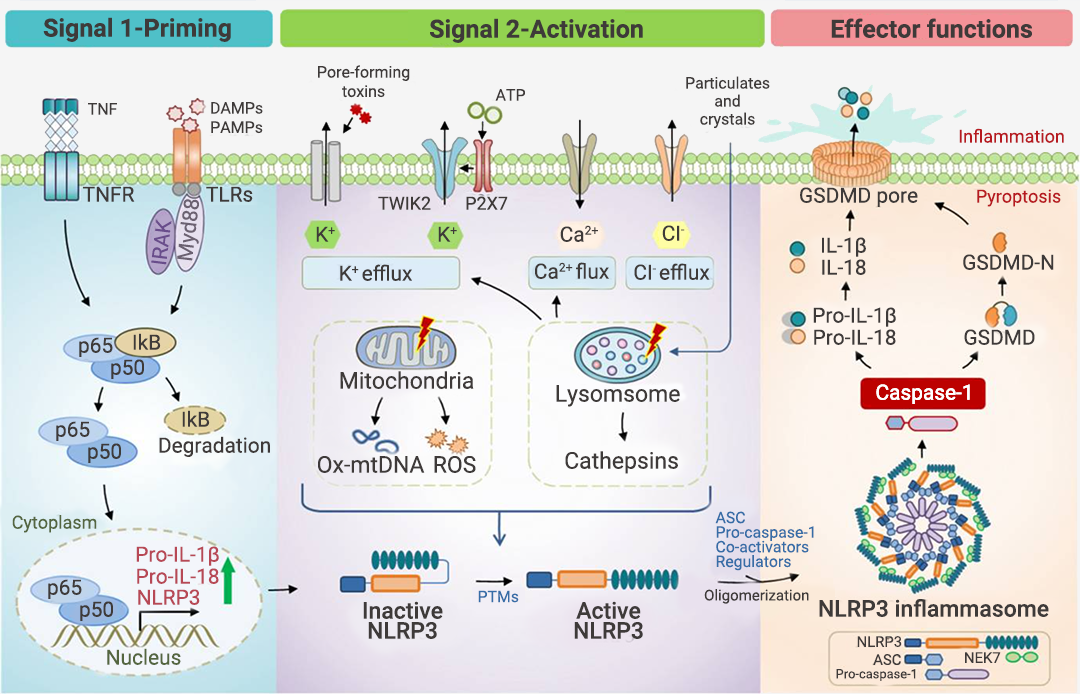
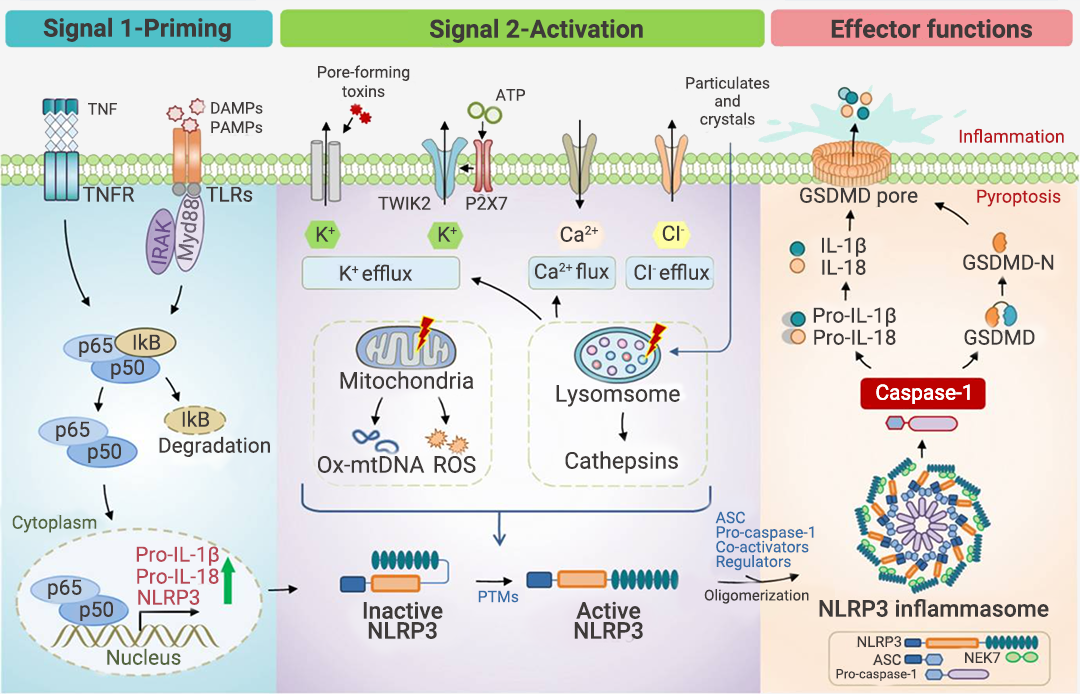
现有研究认为,经典炎症小体通常需要经历启动(通过炎症刺激: 包括 TLR 配体,如 LPS 和细胞因子,如 TNFα)和激活(常见激活源见表 1)两个连续的步骤才能触发炎症小体的组装[8]。NLRP3 炎症小体研究最为广泛,故以此为例(图 2)。图 2. NLRP3 炎症小体的激活[8]。
NLRP3 炎症小体的激活经历三个连续的步骤:
1. 启动过程 (信号 1):包括通过 TLR 识别 PAMP/DAMP 和/或通过 TNFR 感测 TNFα,从而通过 NF-κB 信号传导诱导 NLRP3、pro-IL-1β 和 pro-IL-18 蛋白的表达。
2. 激活过程 (信号 2):包括多种致病性和无菌炎症信号。3. 效应过程:NLRP3 炎症小体的寡聚化和激活导至 Pro-Caspase-1 转变为活性形式 Caspase-1,Caspase-1 进而剪切下游分子 Pro-IL-1β 和 Pro-IL-18 以及 GSDMD (Gasdermin D),从而导至 IL-1β 和 IL-18 的释放以及 N-GSDMD 的形成;N-GSDMD 进入细胞膜,形成孔结构,将成熟的 IL-1β 和 IL-18 释放到细胞外空间,诱导细胞焦亡并引起炎症。
值得注意的是,研究表明,K+ 外流是 NLRP3 激活的必要条件,被激活的 NLRP3 与 NEK7 的结合,也是招募 ASC 和 pro-CASP1 组装 NLRP3 炎症小体的必要步骤[7]。
▐ 非经典炎症小体活化通路
在非经典炎症小体中,Caspase-4,5,11 直接感知细胞内 LPS,随后组装形成大分子复合物[7]。Caspase-4,5,11 通过剪切 GSDMD,从而实现细胞焦亡 (图 3)。图 3. 非经典和经典炎症小体活化通路[7]。 另外,GSDMD 孔隙形成也可导至 K+ 流出,从而导至次级 NLRP3 炎症小体激活和相关细胞因子分泌 (也就是说,非经典炎症小体激活后,能够激活经典炎症小体,从而引发 Caspase-1 裂解 Pro-IL-1β 和 Pro-IL-18,导至机体炎症反应)。

炎性小体介导的炎症与多种人类疾病有很大关系,在急性炎症中,炎症小体的激活有助于去除死细胞并启动组织修复。然而,在慢性炎症中,炎症小体的持续激活是有害的,因为它会损伤组织。
NLRP3 炎症小体的激活主要在单核细胞和巨噬细胞中,也可广泛在内皮、上皮和间质细胞中被激活,因此 NLRP3 炎症小体与皮肤、脑、肝脏等不同器官的多种炎症性疾病相关。
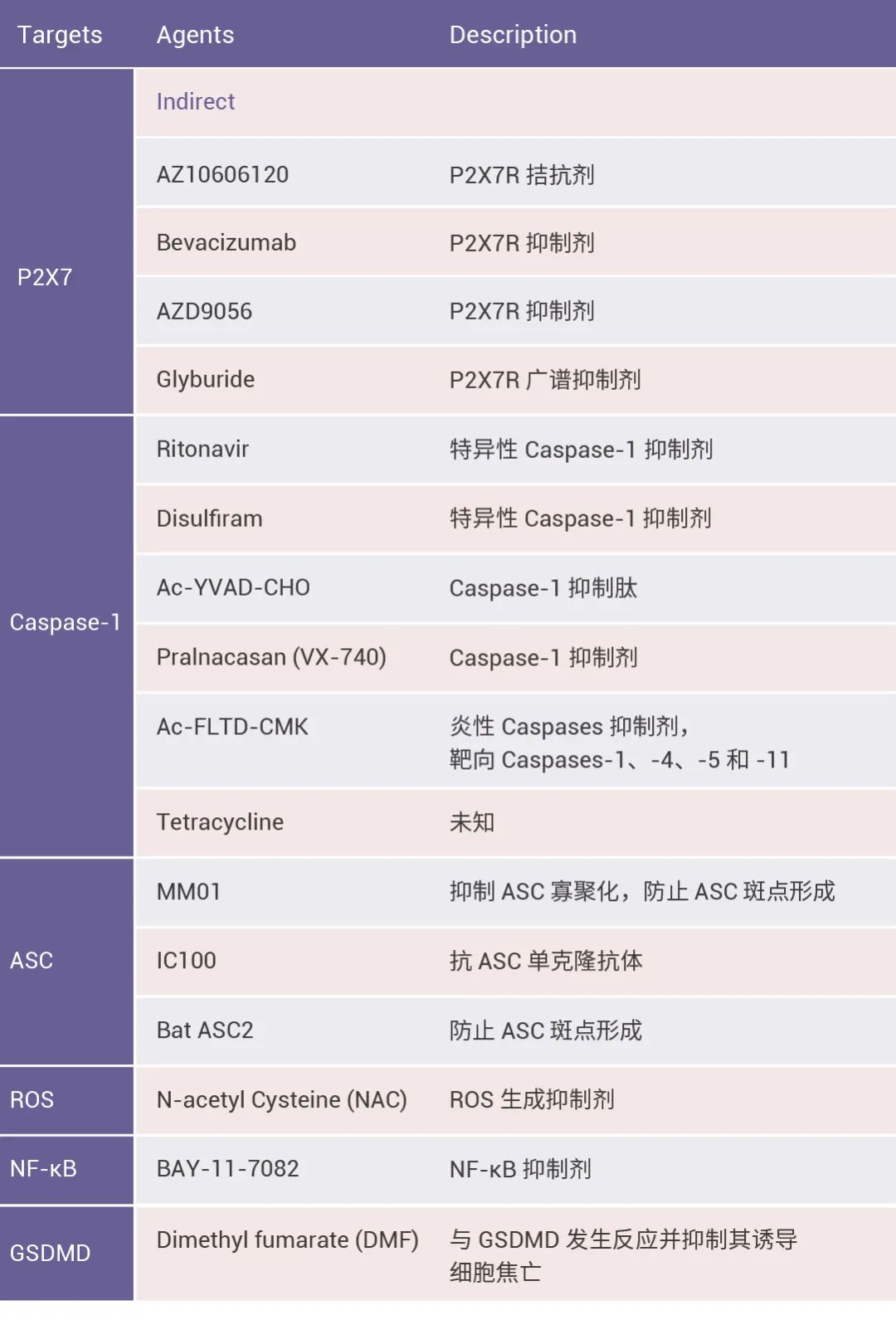
此外,突变诱导的 NLRP3 炎症小体组装激活会引发无菌炎症性疾病,包括家族性寒冷型自身炎症综合征 (Fcas)、Muckle-Wells 综合征 (MWS) 和新生儿多系统炎症性疾病。 图 4. NLRP3 炎症小体机制的药理学靶向[8]。
研究表明,操纵炎症小体的激活状态有利于治疗多种疾病[8]。目前,人们已经开发出多种合成小分子、天然化合物和抗体来靶向炎症小体的组装、炎症小体上下游信号通路以及炎症相关受体 (图 4)。例如,MCC950 通过抑制 NLRP3 炎症小体的激活从而改善了糖尿病肾病小鼠模型的肾脏损伤[9]。
此外,小 M 为大家整理了 NLRP3 炎症小体靶向抑制剂的汇总,大家按需收藏喔~
表 2. 靶向 NLRP3 炎症小体的抑制剂汇总[8][9][10]。 
本篇推文为大家详细介绍了炎症小体的分类 (经典和非经典炎症小体)、炎症小体的活化通路、炎症小体的组装方式、以及炎症小体涉及的人类疾病和 NLRP3 炎症小体靶向试剂的研发进展。这些现有成果是 20 多年来众多研究者一步一脚印所摘取的,但我们所揭露的也只是炎症小体的冰山一角,更广阔的未知犹待你我探索!
MCE的所有产品仅用作科学研究或药证申报,我们不为任何个人用途提供产品和服务。
参考文献:
 [1] Martinon F, et al. The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol Cell. 2002 Aug;10(2):417-26. [1] Martinon F, et al. The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol Cell. 2002 Aug;10(2):417-26.
[2] de Zoete M R, et al. Inflammasomes. Cold Spring Harb Perspect Biol. 2014, 6(12): a016287.
[3] Man S M, et al. Regulation of inflammasome activation. Immunol Rev. 2015, 265(1): 6-21.
[4] Jiang Q, et al. Inflammasome and Its Therapeutic Targeting in Rheumatoid Arthritis. Front Immunol. 2022 Jan 13;12:816839.
[5] Strowig T, et al. Inflammasomes in health and disease. Nature. 2012 Jan 18;481(7381):278-86.
[6] Bergsbaken T, et al. Pyroptosis: host cell death and inflammation. Nat Rev Microbiol. 2009 Feb;7(2):99-109.
[7] Paerewijck O, et al. The human inflammasomes. Mol Aspects Med. 2022 Dec;88:101100.
[8] Chen C, et al. Activation and Pharmacological Regulation of Inflammasomes. Biomolecules. 2022 Jul 20;12(7):1005.
[9] Zhang C, et al. A small molecule inhibitor MCC950 ameliorates kidney injury in diabetic nephropathy by inhibiting NLRP3 inflammasome activation. Diabetes Metab Syndr Obes. 2019 Aug 2;12:1297-1309.
| 
 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 





 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 雷达卡
雷达卡 发表于 2024-2-2 14:21
发表于 2024-2-2 14:21

 提升卡
提升卡


