登陆有奖并可浏览互动!
您需要 登录 才可以下载或查看,没有账号?立即注册


×
铁死亡大家并不陌生,但由于缺乏特定的标记物,在生理条件下识别铁死亡一直具有挑战性。近期,Mol Cell 报道了首个铁死亡的标志物 Hyperoxidized PRDX3 的发现!快随小 M 一起看看吧~
近期,Jin Ye 团队发表的文章 “Identification of hyperoxidized PRDX3 as a ferroptosis marker reveals ferroptotic damage in chronic liver diseases”[1],鉴定出首个铁死亡特异性标记物--Hyperoxidized PRDX3 (过氧化PRDX3)。该研究结果为铁死亡领域提供了一个期待已久的铁死亡标记物,从而能够更精确地检测这种独特的细胞死亡形式。
 缩略词 缩略词
GPX4 : Glutathione peroxidase 4, 谷胱甘肽过氧化物酶 4 SLC7A11: Solute carrier family 7 member 11 , 溶质载体家族 7 成员 11 GSH: Cyst(e)ine-glutathione, 半胱氨酸-谷胱甘肽 GPX4: Glutathione peroxidase 4, 谷胱甘肽过氧化物酶 4 PRDX3: Peroxiredoxin 3, 过氧化过氧化还原蛋白 3 PRDXs: Peroxiredoxins, 过氧化还原蛋白 AFLD: Alcoholic fatty liver diseases, 酒精性脂肪肝 NAFLD: Nonalcoholic fatty liver diseases, 非酒精性脂肪肝 HFD: High fat diet, 高脂饮食

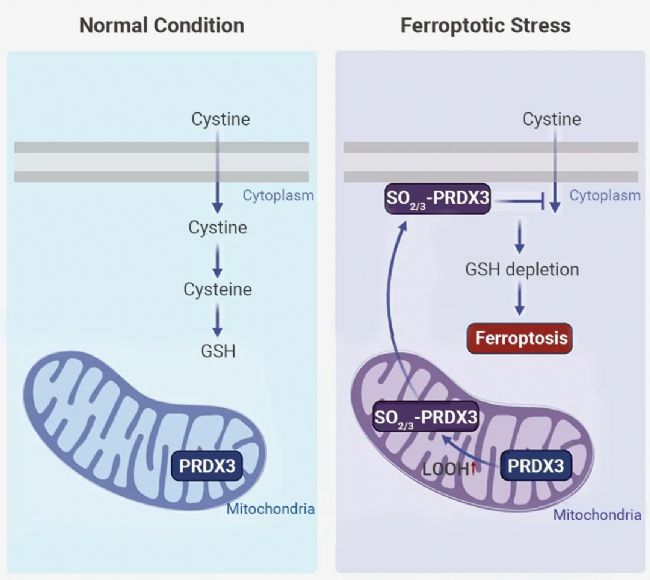
铁死亡是一种由脂质过氧化引起的调节性细胞死亡形式[2]。其主要防御机制是 SLC7A11-GSH-GPX4 轴[3]。在正常情况下 (图 1A),SLC7A11 介导胱氨酸摄取,并产生半胱氨酸 (Cys, Cysteine),促进了 GSH 的生物合成。GSH 是 GPX4 的辅助因子,可有效解毒脂质过氧化,抑制氧化应激损伤。
图 1. 过度氧化的 PRDX3 参与铁死亡[1]。
而在铁死亡胁迫期间(图 1B),PRDX3 由于暴露于线粒体脂质过氧化物而发生过度氧化。随后,过度氧化的 PRDX3 (SO2/3-PRDX3) 从线粒体易位到质膜上 (也可能是线粒体 PRDX3 先重新定位到质膜上,然后在质膜上发生过度氧化),质膜上 SO2/3-PRDX3 的存在抑制了胱氨酸的摄取,从而促进了铁死亡[1][2]。
PRDX3 是一种线粒体特异性过氧化物酶,是抗氧化酶家族 PRDXs 的成员之一。PRDXs 代表分布在各种细胞器中的多种酶抗氧化系统之一,并且根据 Cys 残基的数量和位置对 PRDXs 的不同亚型进行划分。哺乳动物 PRDXs (PRDX 1-6) 的六种亚型具有一或两个保守的 Cys 残基 (Peroxidatic Cys; CP; Resolving Cys; CR)。PRDXs 亚型根据保守的 Cys 残基的数量和位置以及催化循环过程中产生的二硫键的类型分为三种不同的形式。其中最为典型的是 2-Cys PRDX。
在正常情况下,典型的 2-Cys PRDXs,包括 PRDX1-4,可通过催化性 Cys 来还原过氧化物。反应产生的半胱磺酸 (Cys-SOH) 充当连接剂,利用二硫键连接过氧化 PRDXs 产生同二聚体。Cys-SOH 再被还原为 Cys,又可以重新参与到 PRDXs 的催化循环中 (反应 1-3)。 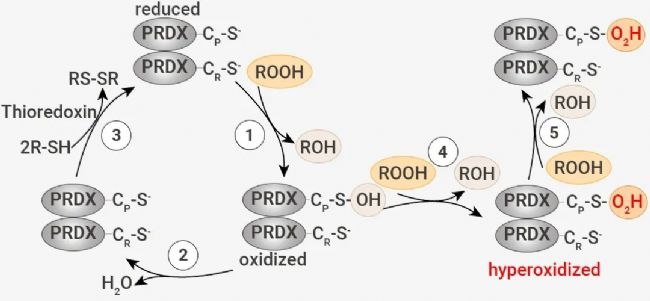 图 2. PRDXs 的过氧化催化循环[1]。 图 2. PRDXs 的过氧化催化循环[1]。当过量的过氧化物积累时,二硫键形成的速率不够快,会导至 PRDXs 中的 Cys-SOH 被过氧化物进一步氧化,生成半胱亚磺酸 (Cys-SO2H)、磺酸 (Cys-SO3H),这是 PRDXs 的过氧化物 (反应 4-5)。因此,脂质过氧化物在铁细胞中的积累可能被 PRDXs 感知,导至蛋白质的过度氧化。 ▐ PRDX3 在铁死亡细胞中被特异性过度氧化
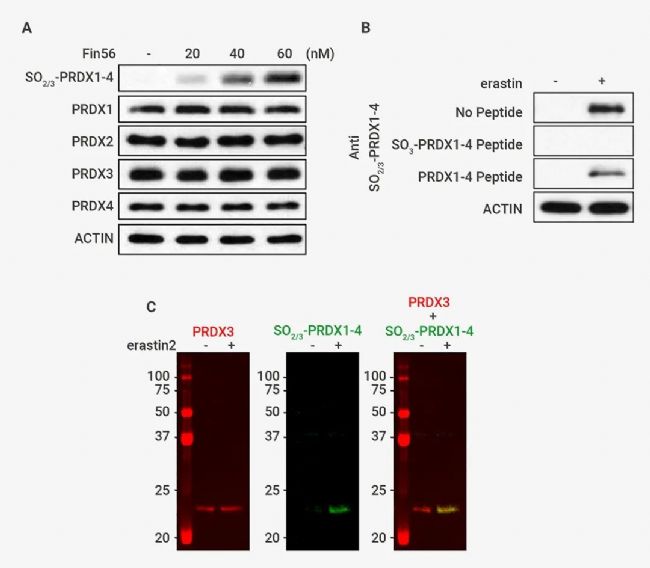
作者使用可识别包含 PRDX1-4 的 Cys-SO2/3 H 的肽的抗体进行了免疫印迹分析。结果发现:(1) 在未经处理的 SV589 细胞中几乎检测不到任何信号; (2) 添加铁死亡诱导剂如 erastin、RSL3 或 FIN56(图中仅显示 erastin) 处理的细胞中观察到~23 kDa 的单条带 (图 3B)。此外,该条带的检测可被抗体与过氧化抗原 (SO3-PRDX1-4 Peptide) 共孵育所阻断,但未被未修饰的对照肽阻断 (图 3B),表明该抗体特异性靶向过氧化 PRDX1-4。
图 3. PRDX3 在铁死亡过程中被过度氧化[1]。 (A) 通过免疫印迹分析测定用指定浓度的 erastin、RSL3 或 FIN56 (此处未显示) 处理12 小时的 SV589 细胞中的过氧化量和总 PRDX1–4 量。(B) 加或不加 2µM erastin 处理 12 小时的细胞,在不存在 2 µg/mL 阻断性高氧化抗原肽或未修饰的对照肽的情况下进行免疫印迹分析。(C) 加或不加 200 nM erastin2 处理 12 小时的细胞裂解物在同一印迹中通过抗 PRDX3 和抗 SO2/3-PRDX1–4 进行免疫印迹,然后用分别红色和绿色荧光标记的二抗进行检测。
由于 PRDX1-4 之间的分子量不同,且仅在铁死亡细胞中检测到 ∼23 kDa 的单个条带,表明在这种条件下只有一个 PRDX 被过度氧化。在同一印迹中通过抗 PRDX3 和抗 SO2/3 -PRDX1–4 进行免疫印迹,由 erastin 衍生物 erastin 2 诱导的过氧化 PRDX 与 PRDX3 一起迁移 (图 3C),证明了 PRDX3 在铁死亡过程中被特异性过度氧化。
▐ 过氧化 PRDX3 通过抑制胱氨酸摄取诱导铁死亡
作者评估了过度氧化的 PRDX3 对其他化合物诱导的铁死亡的影响,结果表明,PRDX3-/- 能够更敏感地抑制直接作用于胱氨酸转运系统的铁死亡诱导剂( 例如 erastin 和 SAS),而对作用于下游 GPX4 水平的诱导剂(例如 RSL3 和 FIN56)敏感度降低。也就是说,过度氧化的 PRDX3 很有可能与 erastin 和 SAS 一样,通过抑制胱氨酸摄取来诱导铁死亡(图 4A)。同时,利用荧光染色定位,研究人员发现本来存在于线粒体的 PRDX3 在过氧化修饰后转移到了细胞膜上(图 4C)。
图 4. 过氧化 PRDX3 可抑制胱氨酸摄取诱导铁死亡[1]。 (A) erastin 处理细胞 24 小时后的活力测定;(B) 加或不加Deferoxamine (DFO, 50 μM) 或 Ferrostatin-1 (Fer, 1 μM) 的情况下,在指定时间内缺乏胱氨酸的指定细胞的活力测定。(C) 加或不加 200 nM erastin2 处理 12 小时细胞,使用指定抗体和 MitoTracker 进行免疫荧光显微镜检查。
随后,将细胞与 erastin 预孵育 9 小时 (9 h 后 WT 细胞中产生过氧化 PRDX3),检查过氧化的 PRDX3 对胱氨酸摄取的影响。结果表明,WT 细胞中的胱氨酸摄取受到抑制,但 PRDX3-/- 细胞中则不受抑制 (图中未显示),也就是说,过氧化的 PRDX3 可抑制胱氨酸摄取。
那么,过氧化的 PRDX3 是否通过抑制胱氨酸摄取来促进铁死亡呢?作者比较了 PRDX3-/- 和 WT 细胞在胱氨酸耗尽培养基中的活力 (该培养基不提供胱氨酸摄取的细胞外来源)。结果表明,WT 细胞中产生了过度氧化的 PRDX3,且可以通过铁死亡抑制剂 Deferoxamine 或 Ferrostatin-1 来挽救 (图 4B)。这些结果表明,过氧化的 PRDX3 通过抑制胱氨酸输入促进铁死亡。
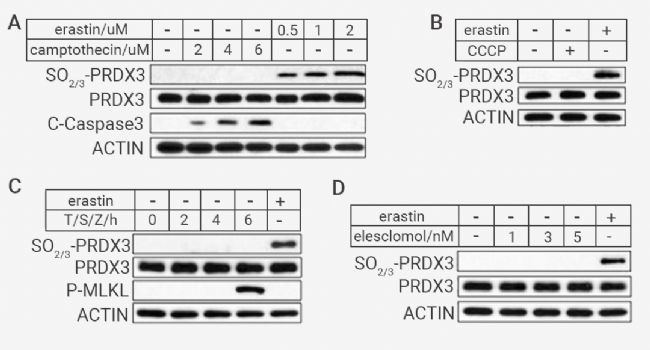
▐ 过氧化的 PRDX3 特异存在于铁死亡细胞中 为进一步评估过度氧化的 PRDX3 是否可以作为铁死亡的标志物。作者发现,在 SV589 细胞中,PRDXs 的过度氧化不因为发生细胞凋亡或者坏死性凋亡而产生 (图 5A, C),也不因为铜死亡诱导剂 Elesclomol 或 CCCP 的处理而产生,PRDX3 只在铁死亡中积累,具有特异性(图 5B,D)。 图 5. 不同细胞死亡类型中,只有铁死亡细胞中存在过氧化 PRDX3[1]。 (A) 通过免疫印迹分析测定用指定浓度的 erastin 或camptothecin 处理 12 小时的 SV589 细胞中过氧化的 PRDX3 和裂解的 C-caspase3 的量; (B) 通过免疫印迹分析测定用 2 μM erastin 或 4 μM CCCP 处理 12 小时的 SV589 细胞中过氧化 PRDX3 的量; (C) 用 20 ng/mL TNF-α、100 nM SM164 和 20 μM z-VAD (T/S/Z) 或 2 μM erastin 处理指定时间的 HT29 细胞中过氧化 PRDX3 和 p-MLKL 的量,通过免疫印迹分析确定; (D) 通过免疫印迹分析测定用 2 μM erastin 或指定浓度的 elesclomol 加2 μM CUCL2 处理 12 小时的 SV589 细胞中过氧化 PRDX3 的量。

作者应用过氧化的 PRDX3 作为标记物,确定了铁死亡是导至 AFLD 和 NAFLD 小鼠模型中肝细胞死亡的原因。过度氧化的 PRDX3 揭示了 AFLD 和 NAFLD 中的铁死亡损伤。在用对照饮食喂养的小鼠中看不到过度氧化的 PRDX3,但在 > 70% 的用乙醇喂养的小鼠(长期和暴饮暴食乙醇形成的 AFLD 小鼠模型)和在约 70% 的饲喂 HFD 的小鼠(长期高脂饮食 (HFD) 喂养的 NAFLD 小鼠模型)的肝脏中检测到铁死亡标记物。此外,在 AFLD 和 NAFLD 小鼠模型中,肝脏中过度氧化 PRDX3 的水平与肝损伤的标志物 AST 和 ALT 水平 (肝损伤标志物) 呈正相关。
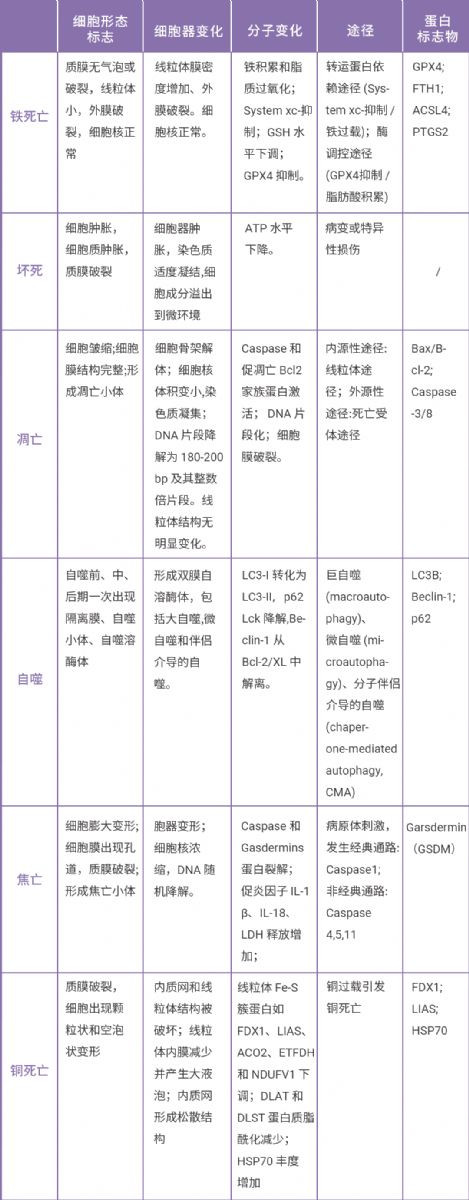
小 M 还为大家整理了相关细胞死亡标志物汇总,有需求的小伙伴可以收藏备用喔~ 表 1. 铁死亡与其他细胞死亡的生物标志物汇总[4][5][6][7]。

本文小 M 带大家一起认识了高分文献中验证 PRDX3 是铁死亡特异性标志物的思路和方法。过度氧化的 PRDX3 通过抑制胱氨酸输入促进铁死亡,决定了它独一无二的身份,为铁死亡研究领域填补了空白的一环。
MCE的所有产品仅用作科学研究或药证申报,我们不为任何个人用途提供产品和服务。
参考文献:
[1] Cui S, et al. Identification of hyperoxidized PRDX3 as a ferroptosis marker reveals ferroptotic damage in chronic liver diseases. Mol Cell. 2023 Nov 2;83(21):3931-3939.e5. [2] Yan Y, Gan B. Hyperoxidized PRDX3 as a specific ferroptosis marker[J]. Life Metabolism, 2023: load042. [3] Li J, et al. Ferroptosis: past, present and future. Cell Death Dis. 2020 Feb 3;11(2):88. [4] Loveless R, Bloomquist R, Teng Y. Pyroptosis at the forefront of anticancer immunity. J Exp Clin Cancer Res. 2021 Aug 24;40(1):264. [5] Frank D, et al. Pyroptosis versus necroptosis: similarities, differences, and crosstalk. Cell Death Differ. 2019 Jan;26(1):99-114. [6] Zhao G, et al. Copper induce zebrafish retinal developmental defects via triggering stresses and apoptosis. Cell Commun Signal. 2020 Mar 14;18(1):45. [7] Wang L, et al. The emerging mechanisms and functions of microautophagy. Nat Rev Mol Cell Biol. 2023 Mar;24(3):186-203.
| 
 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 





 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 雷达卡
雷达卡 发表于 2024-1-16 13:27
发表于 2024-1-16 13:27


 提升卡
提升卡


