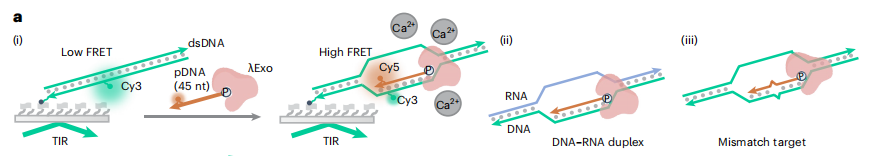
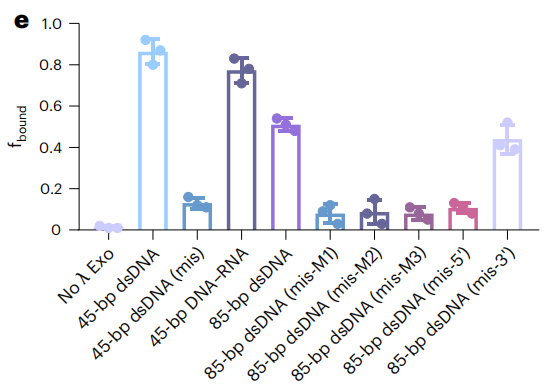
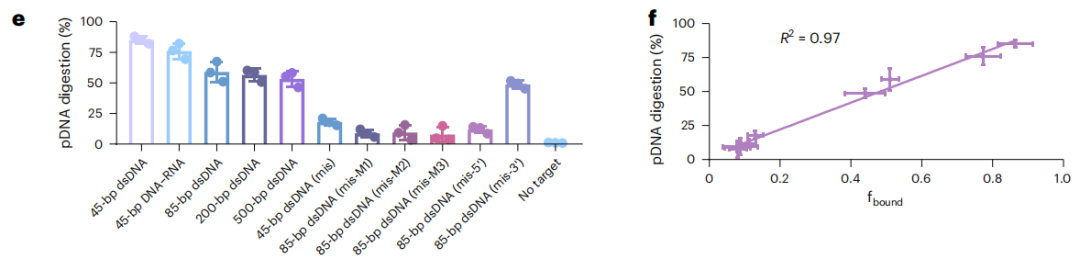
双链核酸的序列特异性识别对于分子诊断和原位成像至关重要。CRISPR系统已经彻底改变了基因组编辑、表达干扰和分子诊断领域。Cas蛋白介导引导RNA(gRNA),通过识别PAM序列来切割目标核酸。此外,在目标识别时,也有一些Cas蛋白对单链报告核酸表现出不加区分的切割活性。然而,CRISPR-Cas系统的PAM依赖性和脱靶效应不仅限制了其目标,但也对识别特异性提出了挑战。这激发了一种对dsDNA进行序列特异性识别的新工具。 近日,杂志Nature biotechnology上发表了一篇题为“Bacteriophage λ exonuclease and a 5′-phosphorylated DNA guide allow PAM-independent targeting of double-stranded nucleic acids”的文章。作者发现了噬菌体λ外切酶(λExo)的酶活性。5 ' -磷酸化的单链DNA (pDNA)可以与dsDNA和DNA-RNA双链上的互补区域结合,λExo在Mg2+存在下会将pDNA消化成核苷酸。该过程对pDNA结合区域内的错配非常敏感,不需要特定的基序。λExo-pDNA可用于检测病原体基因,且识别突变不需要特定的基序,也不需要额外的报告基因,与各种比色或测向流动技术兼容。此外,λExo-pDNA也可用于基因组位点成像。总的来说,作者证明了λExo-pDNA系统是分子诊断和基因成像应用的多功能工具。 图片来源:nature biotechnology 主要内容 λExo-pDNA结合dsDNA的smFRET研究 噬菌体λ外切酶(λExo)识别5’-磷酸化的dsDNA,并降解5’-磷酸化链,留下3’overhang。λExo对非磷酸化DNA和单链DNA(ssDNA)催化活性较低。作者采用TIRFM进行smFRET检测,研究λExo介导的dsDNA结合(下图a)。当dsDNA靶点(45bp的dsDNA)与pDNA长度相同时,观察到显著高FRET状态,表明pDNA在λExo的帮助下与靶点结合,其fbound达到0.86。随着目标核酸长度的增加(85-bp的dsDNA),fbound减少到0.52。当pDNA与DNA-RNA双链靶标的DNA序列互补时,fbound略低于45bp的dsDNA,原因是DNA-RNA双链的稳定性增加。此外,λExo介导的结合对单一错配很敏感。互补区中间和5‘端附近的错配有效地阻碍了pDNA的结合(下图e)。
λExo-pDNA与dsDNA的结合。 图片来源:nature biotechnology 在Mg2+存在下,λExo对pDNA的酶切反应 作者设计了pDNA与cy3标记和cy5标记的链杂交产生荧光信号的结构,形成较高FRET。一旦pDNA链被λExo消化,将导至EFRET值下降(下图a)。在100秒的时间范围内,45bp的dsDNA 分子有75.2%发生EFRET降低;45bp的DNA-RNA双链分子有63.1%发生EFRET降低;85bp的dsDNA分子有42.4%发生EFRET降低;200bp的dsDNA分子有41.7%发生EFRET降低。 pDNA的消化活性对5‘端和互补区中间附近的错配也高度敏感。大多数分子在100 s内没有表现出荧光状态转变。这可能是由于λExo-pDNA与不匹配的靶点的结合亲和力较低。fbound与消化率呈线性关系,说明λExo-pDNA与靶标的结合亲和力决定了催化效率(下图f)。
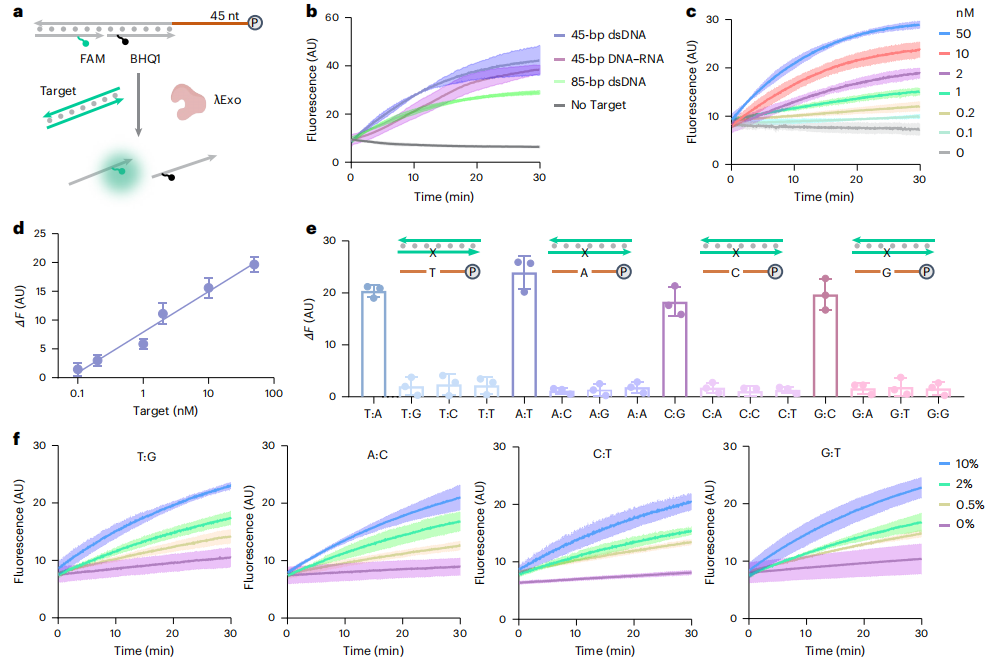
目标核酸在Mg2+存在下诱导pDNA酶降解。 图片来源:nature biotechnology 利用pDNA探针检测ds-核酸 作者开发了一种稳健而快速的方法来检测ds-核酸。将荧光团标记的链和猝灭剂标记的链与pDNA链杂交,导至荧光猝灭。靶标的存在诱导了pDNA的酶促消化,导至荧光的恢复。所有完美匹配的靶标在30 min内都显示出显著的荧光增加(图b)。pDNA探针在浓度低至0.1 nM时即可检测到目标dsDNA(图c,d)。且所有类型的不匹配信号平均比来自匹配目标的信号低10倍(图e)。为了评估低丰度突变体序列的检测能力,作者选择了四种具有代表性的错配类型。在存在许多不匹配序列的情况下,该方法成功地检测到了完全匹配的序列,其丰度为0.5%(图f)。
荧光法λExo-pDNA检测ds-核酸。 图片来源:nature biotechnology 病原体基因的体外检测 提取SARS-CoV-2 RNA,通过逆转录和重组酶聚合酶扩增(RT-RPA)进行扩增。分别设计用FAM和ROX染料标记的pDNA探针检测E和N基因。结果显示每微升1拷贝阳性样本与阴性对照之间E和N基因的荧光信号存在显著差异(图b)。临床阳性拭子样本中这两个基因的荧光明显强于阴性样本(图c)。 作者还证明了λExo-pDNA在分子诊断中的普遍性。ESKAPE病原体、粪肠球菌(ddl基因)、金黄色葡萄球菌(nuc基因)、肺炎克雷伯菌(mdh基因)、鲍曼不动杆菌(gltA基因)、铜绿假单胞菌(oprL基因)和大肠杆菌(dnaJ和ampC基因)。作者设计了针对上述基因片段的pDNA,结果显示λExo-pDNA探针在有靶基因存在的情况下表现出显著的荧光变化,而在没有靶基因存在的情况下没有发现荧光增加(图d)。
病原体的体外检测。图片来源:nature biotechnology 病原体突变型的体外检测 为了证明λExo-pDNA的多重检测能力,采用多路RPA生成dsDNA靶标。用FAM、ROX和Cy5标记3个正交的pDNA,检测S基因的保守序列和编码D614G和N501Y点突变的序列。当pDNA探针被设计成与突变体序列完全匹配时,突变体序列表现出比野生型要高得多的荧光(图g)。从相应的信号强度中可以推断出来自不同突变体和野生型组合的DNA的存在(图h)。这些结果证实了使用pDNA探针与单一酶可以实现多重检测。 总的来说,λExo-pDNA可用于检测病原体基因,且识别突变不需要特定的基序。pDNA可以作为报告基因,不需要额外的报告基因。
病原体突变型的体外检测。 图片来源:nature biotechnology 原位基因成像 荧光原位杂交(FISH)在细胞基因组成像中得到了广泛的应用。λExo-pDNA也可用于基因组位点的原位成像。为了抑制λExo的核酸酶活性,在成像缓冲液中使用Ca2+,而不是Mg2+。使用λExo-pDNA,作者成功地标记了多个端粒位置,与DAPI共定位(图b)。在分析的210个细胞中,端粒数量为49~103个,平均为73个(图c)。λExo-pDNA和标准DNA FISH斑点之间存在高度的共定位(图e)。两种方法鉴定的端粒总数分布完全匹配(图f)。
原位基因成像。图片来源:nature biotechnology 总结与讨论 作者发现了噬菌体λ外切酶(λExo)的酶活性。5 ' -磷酸化的单链DNA (pDNA)可以与dsDNA和DNA-RNA双链上的互补区域结合,而不需要PAM样基序。结合后,λExo-pDNA系统在Mg2+存在下催化将pDNA消化成核苷酸。该过程对pDNA结合区域内的错配非常敏感,从而在各种应用中具有特殊的序列特异性和减少脱靶效应。不需要特定的基序,如PAM序列,极大地拓宽了靶标的范围。λExo-pDNA可用于检测病原体基因,且识别突变不需要特定的基序,也不需要额外的报告基因,与各种比色或测向流动技术兼容。此外,λExo-pDNA也可用于基因组位点成像。总的来说,作者证明了λExo-pDNA系统是分子诊断,DNA计算和基因成像应用的多功能工具。 此技术未来的发展方向。首先,由于pDNA探针设计的灵活性,通过一次λExo周转可以释放多个报告链。因此,可以进一步增强核酸检测的荧光信号强度。其次,可通过优化pDNA的长度,或开发λExo的变体,进一步提高对单核苷酸错配的选择性,用于检测临床样本中的低频突变,如循环肿瘤DNA。第三,优化实验组分和浓度,有助于实现一锅反应。最后,开发λExo变体,保留目标结合活性,同时消除核酸酶活性,从而能够在活细胞中进行成像。 |

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号