1、标签,杂交和PCR产物的识别和检测 经典的透射电子显微镜已经将杂交可视化(图1)。
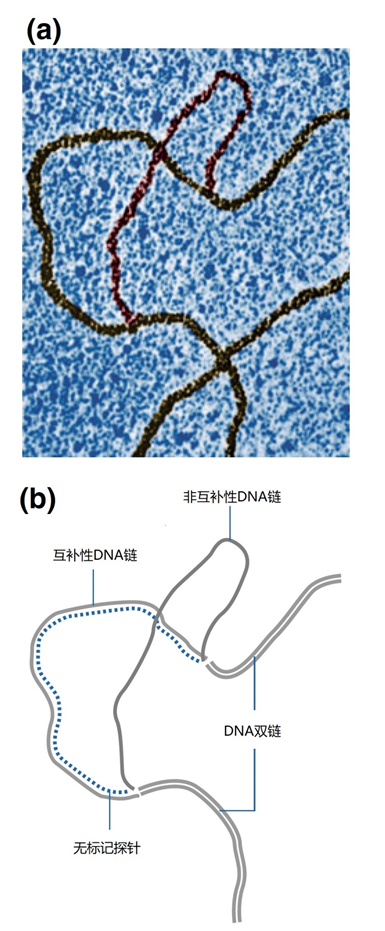
图1 | 基因组DNA杂交的可视化,通过透射电子显微镜(TEM)和阴影技术获得 对单链和双链DNA的区分是基于分子的厚度。很明显,探针已经取代了原生DNA。 它可以清楚地看到探针是如何取代原来的互补链的。目前,杂交完全是通过标签来观察的,如果有必要,还可以通过报告基团来观察(见图2)。
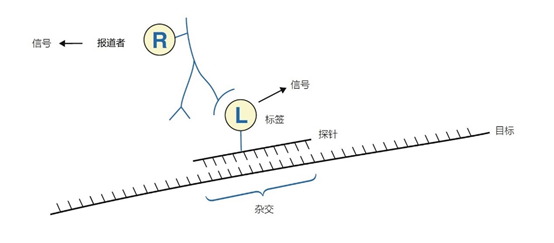
图2 | 杂交和杂交探针检测的术语 标记的探针与单链靶点的互补序列杂交,存在于开口线圈形成中。标签本身可以产生信号,如荧光色素。如果标签不产生信号,可以在标签上附加一个报告基团。报告基团的一个例子是标签的特异性抗体,如地高辛(DIG)标签可以连接到报道碱性磷酸酶和抗DIG抗体。通常情况下,需要进行酶促检测反应。为了检测,标签生物素也可以直接与报告基团、链霉菌素偶联 不同类型的标签通过间隔物与尿嘧啶共价结合,可以纳入探针中。已经证明,间隔物的最佳长度在11到16个原子之间。间隔物使标签与新形成的杂交体有一定的距离,从而避免了立体阻碍,不仅在标签和探针与靶点的杂交之间,而且在标签和报告基团之间(如特异性抗体与合子的结合)(图3)。
图3 | 从PCR中捕获标记扩增产物的两种方法 a 基于互补序列,标记的扩增物将被固定在固相(如玻璃或尼龙膜)上的探针捕获。除非标签是荧光色素,否则将使用判读器进行检测。 b 生物素化的PCR产物与序列特异性标记的PCR产物一起被捕获在固相上,用链霉亲和素涂布,捕获后用报告基团检测 这样的改性核苷酸被称为核苷酸类似物。著名的例子是生物素-16-(d)UTP和地高辛-11-(d)UTP,其中数字16和11表示构成间隔物的原子数。C-原子是最常用的原子。标签还可用于捕获扩增物,例如从PCR混合物中捕获扩增物,并将其结合到载体上,如ELISA板、X-MAP®技术中的珠子或尼龙膜上。因此,标签具有配体的功能(见图4)。
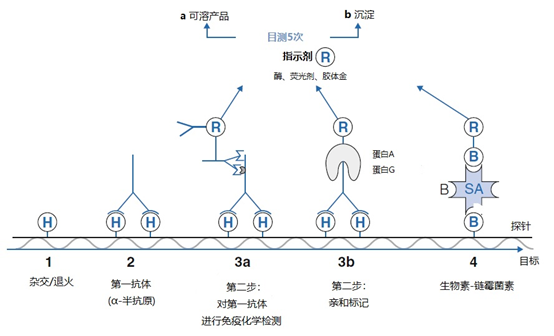
图4 | 使用免疫化学和SABC方法的标签检测系统 在免疫过程中,标签H作为一个合体物(1)。半抗原特异性多克隆和单克隆抗体与标签结合(2)。随后的步骤可以有所不同。指示剂(R)和二级抗体(3a)之间的结合物提供了通过酶的活性、荧光或银增强产生信号的机会。同样提供报告基团的蛋白质A和G,对抗体的Fc片段有很高的亲和力(3b)。生物素对链霉菌素有极高的亲和力,报告基团可以通过生物素进行偶联(4)。检测的最后一步通常是酶促反应,产生可检测的信号(5)。在步骤5之后可以引入一个信号放大的步骤。用光谱法测量的可溶性产物或可目测或用显微镜检查的沉淀物 2、加入标签 标签对于许多分子诊断测定是必不可少的。标签可以通过不同的方式纳入引物和探针中,化学和酶法是最经常使用的方法。 2.1、化学合成 寡核苷酸可以在化学合成过程中被标记。一个或多个核苷酸在必要的位置被带有标签的核苷酸类似物取代,并具有正常的沃森和克里克碱基配对。 2.2、化学标签 铂金与ssDNA和ssRNA结合的亲和力被用来与通用连接系统(ULS)以通用的方式耦合标签(图5)。
图5 | ULS标记 咪唑环中的C2原子被铂金和一个带有半抗原或荧光色素的间隔物所取代。铂对胍的N7原子的亲和力非常高,会发生稳定的结合。在85℃下孵化30分钟后,单链DNA或RNA分子可以被标记。核苷酸处于一个开放的线圈形成中。孵育步骤后,通过在冰上直接冷却,分子将保持单链形成。一个纯化步骤将结束标记反应。 另外,活化的小分子(触媒、荧光剂或功能性酶片段)可以与修饰的氮基进行共价偶联。 2.3、酶标法 酶法用于合成大于30个核苷酸的标记探针。这是一个两步程序。首先,生产大量的DNA或RNA。随后,用酶法合成DNA或RNA来加入标签。部分dNTPs被标记的核苷酸取代,如DIG-11-dUTP或生物素-11-dUTP(见图6),从而得到一个标记的探针。
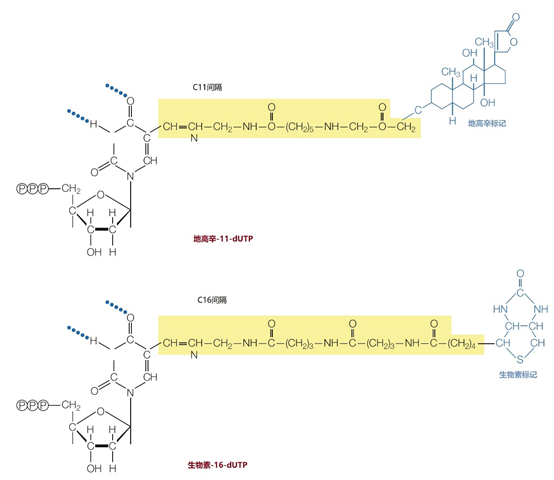
图6 | 常用的标记核苷酸以及地高辛-11-dUTP和生物素-16-UTP的结构式 这些标签通过间隔物分子与尿嘧啶的5′原子共价连接。数字11和16表示两个分子之间间隔物的长度。间隔物使标签与杂交探针有足够的距离,以避免在后续步骤(标签的可视化)中出现立体阻碍。这两种标签都可以用特定的抗体进行检测。生物素也可以用链霉菌素检测。虚线表示尿嘧啶分子上可能产生氢键的地方。除了地高辛和生物素外,荧光色素也可以与核苷酸偶联。 自过去20年以来,切口平移和「随机引物标记」已被频繁使用。 2.4、切口平移 在切口平移策略中,dsDNA将与两种酶,即DNase-I(外切酶)和DNA依赖性DNA聚合酶(图7 a)以最佳比例进行孵化。DNase-I切割5′-3′共价键,并在每条DNA链的随机位置形成缺口。该缺口现在呈现出一个自由的3′-OH分子,是新的DNA合成的起点。在上游的核苷酸被5′-3′-解核酸酶活性去除后,一个新的核苷酸将通过3′-5′-DNA合成取代它。缺口沿着链子移动,当它遇到(相反的)反平行链上的缺口(不连续)时就会停止。探针的片段大小取决于缺口的数量,通常为400bp。
图7 | 酶法探针标记方法 a 切口平移:dsDNA与外切酶和DNA依赖性DNA聚合酶(大肠杆菌DNA聚合酶I)一起孵化。DNA聚合酶I会破坏5′-3′-共价键,导至每条DNA链上出现缺口。每个缺口的自由3′-OH端将作为DNA聚合酶I的连接点,它在5′-3′方向上开始新的聚合。已经存在于链上的核苷酸将被5′-3′-解核酸酶域去除。另一条链则作为基质。DNA聚合酶I没有能力在5′端进行连接。因此,破口从3′向5′移动,当酶遇到反平行链上的下一个破口时,DNA的合成将停止。探针的片段大小(平均为400bp)取决于破口的数量。在DNA合成过程中,标签将通过标记的核苷酸取代部分dNTPs而被纳入。 b 随机引物延伸。双链DNA将被变性,单链分子与具有随机序列的六核苷酸混合物孵化。这些六核苷酸退火到ssDNA的互补区域,并作为DNA聚合的起始点发挥作用。所有这些退火的引物都是DNA聚合酶I的Klenow片段的起点。一定比例的标记的类似核苷酸序列如DIG-dUTP或生物素-dUTP可用于合成探针。探针序列的长度是不同的。六聚体的密度越高,片段越小 2.5、「随机」引物标记 双链DNA的变性是「随机引物标记」的第一步(图7 b)。两条单链同时作为使用大肠杆菌的DNA聚合酶I的Klenow片段进行DNA合成的模板(见图8)。
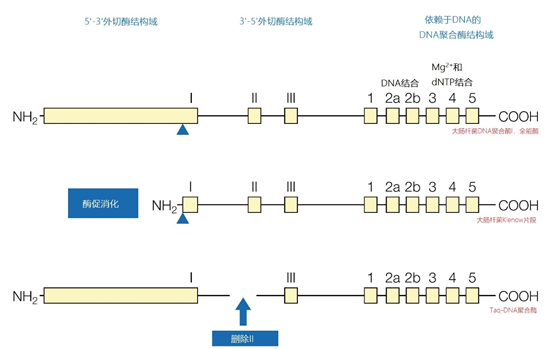
图8 | 不同DNA聚合酶之间的结构同源性 与大肠杆菌的全酶DNA依赖性DNA聚合酶I相比,Taq-DNA聚合酶由于3′-5′-外切酶结构域的第二外显子缺失而缺乏校对特性。Klenow片段是通过对DNA聚合酶I进行酶促处理得到的。Stoffel片段是一种重组的Taq-DNA聚合酶,具有校对功能,但缺少5′-3′-外切酶结构域。 然而,依赖DNA的DNA聚合酶需要一个带有3′-羟基末端的短双链DNA片段。否则,它不能开始DNA的合成。合成的具有随机序列的六核苷酸引物在六核苷酸引物的许多部位与变性的单链DNA杂交。聚合酶从3′-OH端拉长短的六核苷酸片段。以一种依赖浓度的方式,杂交的六核苷酸越多;起始点就越多,片段就越短。 2.6、终端标签 有多种末端标记的选择。特殊的酶会在寡核苷酸的3′或5′末端位置上配备一个或多个标签。末端标记主要用于合成的寡核苷酸探针和PCR产品,并且大多使用氟色素作为标签。 |

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号