在前面的系列文章中,我们介绍了细菌、古细菌、病毒和真核生物的DNA情况,今天,我们将介绍它们的RNA。 RNA是一种单链多核苷酸,可以区分为几种形式,每种形式在基因的表达中都有自己的功能。 所有的RNA分子都参与了DNA的转录和翻译成多肽链的过程。 RNA的主要类别是tRNA、hnRNA(前mRNA)、mRNA和rRNA。哺乳动物细胞(2.5×106)含有10 μg RNA,其中80%以上是rRNA。 在这样一个样品中,大约有106~107个mRNA分子存在(图1)。最后,低分子部分还包括ncRNA、snRNA、miRNA和siRNA。
*总RNA和poly(A)-RNA的数量取决于细胞的类型、组织和生理条件。 图1|RNA类型的产量和分布 粗略估计从2.5×106个真核细胞中分离和纯化的RNA类型、产量及其分布。mRNA是指通过使用基于poly dT的亲和层析法可以分离的部分。 单链核酸分子的重要结构是所谓的发夹环(图2a)。这些结构是由分子内碱基配对形成的,通过这种配对,双链和单链环状部分交替出现。
图2|由成对和未成对片段组成的单链RNA分子的示意图 a. 茎环结构(发夹)由互补的富含GC的片段组成。这个基础有一个螺旋结构,而单链环常常堆积在螺旋上。RNA分子具有三维结构,因为有成对和未成对的片段组合。 b,在此基础上可以耦合一些功能特性。 如果链上含有主要由G-C碱基对组成的发夹环,这种结构元素非常稳定,可以对生化反应产生负面影响,如化学修饰、复制、PCR和测序。这种现象被称为碱基压缩。每个RNA分子都有一个独特的三维结构,允许与某些蛋白质进行非常特殊的相互作用(图2b)。 基因表达始于互补的RNA链的合成,基因组DNA起着基体的作用。这个过程被称为转录。作为RNA基质的DNA链被称为反义链(或非编码链或-链)。另一条DNA链(有义链,编码链或+链)的碱基顺序与合成的RNA相同。这种RNA也被称为转录本(表1)。
表1|真核生物中DNA和RNA转录方面最重要的概念概述* 备注 *在细菌中,两条链都将用于转录,这与真核生物相反。这意味着本表中的术语(如有义和反义)可以互换,并与转录本有关。 **除非有义链上有起始密码子,否则可能会从一个完全不同的起始位置产生一个完全不同的转录本。这条链将成为反义链。这种功能在真核生物和原核生物中都可以找到。目前还不清楚这是否会导至蛋白质产物或siRNA或miRNA。 ***为研究目的进行排序的通常程序是从反义链开始的。在诊断中,两条链都将被测序:从反义链开始正向测序,从义链开始反向测序。 在真核生物中,这一过程发生在细胞核内。不同的初级转录本,即异质核糖核酸(hnRNA或pre-mRNA),在真核细胞核中发现,大约有10,000个拷贝。 hnRNA含有非编码序列,称为内含子,在核内通过剪接去除,剪接后,编码外显子被连接到最终的信使RNA(mRNA)。 mRNA被运送到细胞质中,34个三联体编码被用于翻译,多肽的合成过程(图3和4)。
图3|将初级转录本修饰成功能性真核生物信使RNA 基因由外显子(编码多肽链(蛋白质))和内含子(中间序列)组成。后者将在一个称为剪接的过程中从初级转录本(hnRNA,pre-mRNA)中去除。5′端装有7-甲基鸟苷三磷酸(m7GPPP),而3′端将通过一个酶的过程(聚腺苷酸化)装有一个多A尾。第一个和最后一个外显子不是完全的编码区;这些是带有起始(ATG)或终止(TAG、TAA、TGA)密码子的非翻译区(UTRs)。基因三联体编码,合成多肽链,将通过mRNA链的翻译在核糖体中发生。 通过交替剪接,可以从一个基因产生多种蛋白质。这一过程在真核生物中经常发生,在人类和玉米中都有报道。在古细菌中发现了一种独特的拼接形式。 在原核生物中,不具备细胞核,不存在hnRNA的步骤。没有内含子的mRNA直接由基因组DNA产生,不发生拼接。 在tRNA中已经报道了「自我拼接」,这是原核生物和真核生物的共同特征。 原核生物的mRNA转录物包含多个基因(一个所谓的操作子)。这被分割成不同的功能、蛋白质编码序列,大多属于同一代谢途径。这些多克隆mRNA比一般的单克隆(真核生物)mRNA大得多,后者只编码一条多肽链。 1 // 信使RNA (mRNA) 所有细胞RNA中只有相对较小的一部分(约3%)由mRNA组成。一个平均的mRNA约为20~40kb,包含各种功能单元或结构域。 大多数真核生物的mRNA在3′端有一个poly-A-tail,在5′端有一个甲基化的胍(所谓的帽结构)。 3′和5′两端的各种区域不被翻译成蛋白质,即非翻译区(或UTRs)。然而,它们确实在调节蛋白质合成方面发挥了很大的作用。mRNA的区域,翻译成蛋白质,被称为开放阅读框架(ORF)。 细胞核糖核酸不断分解RNA,因此,一个mRNA分子的半衰期一般是几分钟到几小时。 并非一个ORF中的所有基因都被读取。信号分子(如蛋白质和RNA)在其中发挥了重要作用。 此外,DNA在特定区域的甲基化,如调节位点,可以阻止一个基因的转录。在一个转录活跃的细胞中,大约有1%的基因组被读取。许多mRNA,分布在整个细胞质中,以低拷贝数(1~15拷贝)出现。 只有几个基因是高度表达的,在一个给定的细胞内有107~108份mRNA。细胞类型、细胞周期的阶段、发育状态、病理和/或对内部(激素)或外部刺激的反应对细胞中存在的mRNA的性质和数量有很大影响。 2 // 转移核糖核酸(tRNA) tRNA大量存在于细胞核和细胞质中,它不与蛋白质相联系。tRNA的三维结构显示出很大的相似性,但它们的独特之处在于通过特定的反密码子三联体动机来结合一种氨基酸的能力。 3 // 核糖体RNA(rRNA) 在核糖体中(图4),可以发现rRNA,特别是与大约40种核糖体蛋白相关联。
图4|对真核生物80S核糖体的整体结构的印象 40S和60S亚单位以及不同的rRNA(核糖体RNA)分子被标出。此外,图中还描述了mRNA和「成长中的」多肽链的位置(每个氨基酸用一个球体表示)。 核糖体存在于细胞质和有自己的蛋白质合成的细胞器中,如线粒体和叶绿体。核糖体由两个亚单位组成:一个大亚单位和一个小亚单位,都含有rRNA(表2)。
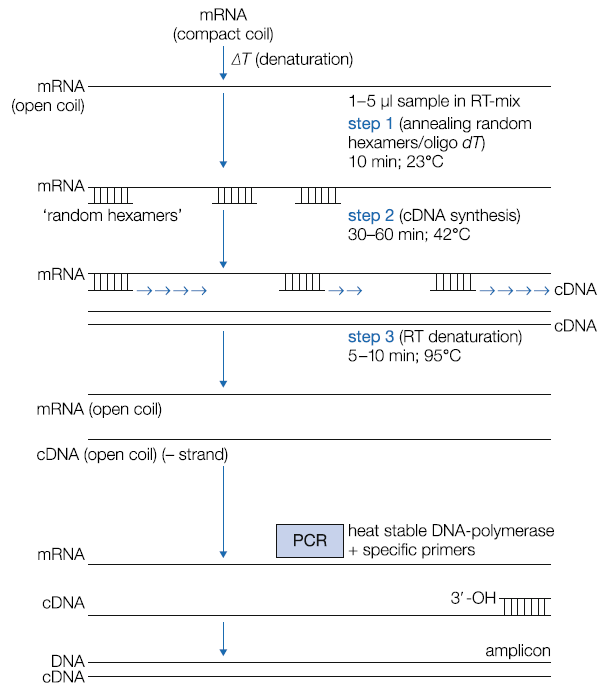
表2|原核生物和真核生物中不同核糖体亚单位的特性(分类见表2.1),与大小和额外的rRNA有关 注释 真核生物的线粒体核糖体与细菌的核糖体之间存在着进化关系 *内共生理论是指线粒体和叶绿体等细胞器起源于自由生活的细菌,在进化过程中被转移到真核细胞中,并部分利用其自身的基因、核糖体和相关的蛋白质合成成为功能实体。 **斯维德伯格常数(S)是衡量分子在离心后的沉淀程度,取决于物体的特性(结构、重量)和分子所处的基质。 ***线粒体的rRNA含量;这些核糖体比其他核糖体小很多。 ****除植物外,5S线粒体rRNA是由基因组DNA编码,而不是由mtDNA编码。这种分子将通过一种转运蛋白从细胞质中转移出来。 由于富含GC的互补序列的存在,每个rRNA都有自己的三维组织。 rRNA位于核糖体的中间,与真核生物的核糖体相比,古细菌、细菌、线粒体和叶绿体的核糖体在亚单位和各种rRNA的大小上更为相似(表2)。 rRNA在蛋白质的合成(翻译)中起作用。内部碱基配对形成功能域。这些强烈保守的结构域在mRNA和tRNA/氨基酸复合物之间的相互作用中至关重要,有利于新生肽链中肽键的形成。 并非所有的rRNA都是高度保守的类型。科、(亚)种和基因型之间的序列差异可用于分型和分类。这种分型主要用于细菌和真菌的rRNA上的物种序列,可以直接或通过PCR间接地进行。 内部转录间隔物(ITSs)是位于编码rRNA的rDNA基因内的特殊DNA序列。在一个rDNA基因中编码有三种rRNA,在细菌中是5′-16S-23S-5S-3′操作子,在真核生物中是5′- 18S-5.8S-28S-3′基因。 rDNA基因在细菌中以有限的数量分散在基因组中(如大肠杆菌中的7个rDNA操作子),在所谓的串联重复中以300~400个拷贝一个接一个分散在5条染色体上(真核生物)。真核生物的rDNA编码单元由非转录或基因间间隔物间隔,这些间隔物划定了单个rRNA基因重复的5′和3′末端。 在单个rRNA基因内,分别在细菌和真核生物的16S/18S编码区的上游发现一个外部转录间隔体(ETS),一个内部转录间隔物(ITS),对细菌和真核生物来说是相似的,分别位于16和23S以及18S~5.8S编码区之间。 此外,真核生物拥有第二个ITS(ITS2),位于26/5.8S基因之间,但在细菌中没有发现类似的序列。作为转录的第一步,整个rRNA基因被转录成hnRNA。然后ITS通过拼接被切除,不构成纳入核糖体的功能性rRNA分子的一部分。ITS的一部分被进一步加工成两个tRNA。 ITS在进化过程中非常保守,并被用于基因分型,如通过基因组ITS DNA的特征鉴定细菌和真菌。它也非常适用于系统发育分析。 4 // 非编码RNAs 以前人们认为不同大小的非蛋白编码RNA(ncRNA)是剪接的副产品,没有生物功能。 1993年,人们清楚地认识到microRNA(miRNA或miRs)和small interfering RNA(siRNA)是有功能的分子。 miRNA的长度约为22个核苷酸,在基因组的各个部分的有义和反义链上都能读到。miRNA在翻译和转录的调节中非常重要,它也与癌症等疾病有关。 小核糖核酸(snoRNA)在rRNA的成熟过程中发挥重要作用,通过甲基化和将尿苷转化为假尿苷。 2012年,两类ncRNA被区分出来,即结构性的和调节性的。 tRNAs、rRNAs、snRNAs(在剪接体中)和snoRNA属于第一类,具有重要的功能作用。基因调控由ncRNAs控制,如siRNA、piRNA、miRNA、lncRNA和长基因间ncRNA。 特殊类型的miRNA和ncRNA(例如,小的和长的非编码RNA)在癌症患者的血清中被发现,数量可观,可以量化。这样,miRNA和lncRNA可以越来越多地被用作生物标志物来指示恶性肿瘤或与特定类型的恶性肿瘤有关。 大约40%的miRNA基因位于基因的内含子上。由于内部碱基配对,许多类型不会被RN酶降解。 除真菌和藻类外,所有真核生物都有miRNA。siRNA在进化过程中非常保守,甚至在古细菌中也有。siRNA作为互补链与功能性mRNA中的关键位置结合,起到沉默mRNA的作用,因此得名。 5 // RNA和聚合酶链反应(cDNA) RNA分子是分子诊断中的一个重要目标,但snRNA、tRNA和hnRNA除外,聚合酶链反应只能以双链DNA为模板进行。因此,有必要合成rRNA或mRNA的DNA拷贝,为通过PCR扩增做准备。源自第六类逆转录病毒的酶(逆转录酶)可以将mRNA和rRNA转录成拷贝DNA或cDNA(图5)。
图5 从RNA制备cDNA 在(无RNase)分离出RNA后,RNA将被热变性,随后在冰上冷却,以解开所有稳定的发夹环,并获得开环的RNA结构。在适当的条件下,引物(随机六聚体、基因特异性引物或oligo(dT))将与RNA的互补区退火,目的是获得具有3′-OH末端的短ds结构。随后,逆转录酶(一种以RNA为基质合成DNA的酶)会合成cDNA。病毒酶确实是在42℃下合成的;热稳定的DNA聚合酶形式,如Thermus thermus,在更高的温度下有活性。这个过程可以在一个温度下进行,在这个温度下,稳定的毛细管将保持开放的线圈结构,引物保持杂交状态。合成的cDNA可以直接作为PCR检测的样本。另一个选择是通过加热使cDNA变性,以使逆转录酶失活,之后可以将其储存在-20℃的环境中。 也可以使用某些具有RT特性的热稳定DNA聚合酶来合成cDNA。 Tth DNA聚合酶常被用于此目的,它的额外好处是对某些血液成分的抑制不敏感。这最后一类酶的主要优点之一是cDNA的合成可以在如此高的温度下进行,具有稳定发夹环和碱基压缩的区域是单链的。 这使得RNA模板可以被寡核苷酸引物所接触。酶在引物与RNA结合的部位开始DNA合成,形成一个短的双链片段。 逆转录酶识别成对的3′-OH末端,并以RNA为模板合成一条新的互补DNA链。有三种类型的引物可供选择:随机寡聚物(主要是六聚物),它与RNA上的许多互补位点结合;识别保守序列的(基因)特异性引物和聚dT引物,它只适用于含有聚A尾的真核生物mRNA。 哪种类型的引物适合,要通过实验来确定。这些引物的混合物也是存在的,含有寡头(dT)和随机六聚体。 在图5中,随机六聚体被作为一个例子使用。 在反应之后,cDNA的混合物被特异性地合成,反转录酶被热变性,mRNA/cDNA被融化。在反应管中加入额外的PCR缓冲液、特定的引物和Taq DNA聚合酶。只有形成被PCR酶识别的双链DNA片段的cDNA结合的PCR引物才会被扩增。 好了,以上就是今天分享的全部内容,如果大家有什么疑问,欢迎在后台给我们留言,或者加入我们一起讨论~ |

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号