合成生物学作为迅速发展的交叉学科,已在生物医药、新材料、诊断、能源和数据储存等领域展现越来越广泛的应用潜力。合成基因组学作为合成生物学的重要研究方向,着重于新生命体系的从头设计与合成,其以Oligo(寡核苷酸链)合成、拼装和转移等核心技术为支撑。新生命体系的从头设计与合成不仅需要合成组成基因组的小片段DNA片段,还需要通过后续的组装与拼接获取完整的合成基因组。Oligo合成及基因组DNA拼装、转移技术是合成基因组学乃至整个合成生物学领域的核心技术体系,今天主要来介绍体外单链DNA(Oligo)合成与双链DNA拼接组装技术。 1.1 化学法 Oligo的化学合成研究始于20世纪50年代,Michelson和Todd首次报道采用磷酸二酯法实现了寡聚二核苷酸的合成。到20世纪80年代,Beaucage和 Caruthers开发了基于亚磷酰胺的DNA合成方,即今天Oligo自动化生产所采用的主要方法。该方法包括去保护、偶联、加帽(可选)及氧化 4 个步骤 (图 1)。由于随着链延长所带来的化学反应效率、合成纯度以及产率的下降,目前该方法合成的Oligo长度一般不会超过200个核苷酸(nt)。为了提高通量,从20世 纪90年代开始发展起来的基于微阵列芯片的DNA合成策略,使得合成成本降低了若干个数量级。然而由于芯片特有的不均一性及边缘效应等原因,相较传统的柱法合成,芯片法合成在长度、准确性方面均有所下降。为了增加芯片合成的精确度,科学家采用不同的技术策略,从各异的芯片产物中挑选出正确合成的寡核苷酸原料。Kosuri采用了多重PCR策略,选择性扩增目的寡核苷酸片段,结合酶促纠错等方法,可以合成长度超过200 nt的寡核苷酸原料,实现了更大规模和更高精度的合成。
图 1.固相亚磷酰胺法从头合成寡聚核苷酸链的4步反应 4步反应包括:(1)去保护。酸催化去除 DMT(二甲氧基三苯基甲基)基团,以便下一轮碱基(dA、dC、dG和dT)添加。(2)碱基偶联。将含有DMT保护基团的亚磷酰胺通过四唑活化剂加到未保护的5′OH末端。(3)加帽。将游离的5′OH乙酰化,以防止进一步的链延伸所造成的单碱基缺失。(4)氧化。通过碘液将磷酸三酯氧化为磷酸盐,进入一个反应循环. 1.2 酶促从头合成法 目前亚磷酰胺法是商业化Oligo合成的主流方法,但其合成的长度限制在200nt左右,而且合成过程中亦会大量使用有毒化学试剂。由于在合成准确性及不需要使用毒性化合物等方面的潜在优势,酶促合成方法受到了越来越多的关注。与化学法相比,酶促法的作用条件温和,对DNA的损伤较少,有助于准确性的提高,同时减少了副产物的产生,可实现更长Oligo的合成。然而,酶促法的发展缓慢,至今未能实现商业化。根据Jensen和Davis对DNA酶促从头合成发展的总结,末端脱氧核苷酰转移酶(TdT)介导的酶促合成法是较好的选择,但还需要更进一步优化。TdT介导的DNA合成技术开发中首先需要解决单个dNTP添加后的终止效率及末端重新活化的问题。近期,Palluk等提出了一种解决方案,他们将 dNTP通过可光诱导剪切的连接子与TdT连接,所得dNTP-TdT复合物可在10—20s的时间完成DNA链的延伸,而且可以重复进行,实现特定DNA链的合成。该方法将为具有实际应用价值的酶促DNA合成法的开发打下基础。 2.1 Gibson 组装 Gibson组装最早由Gibson等在2009年提出,原理如图2所示。该技术基于5′核酸外切酶、DNA 聚合酶及连接酶,利用DNA片段的两端同源序列(其长度通常为15−40bp),实现DNA组装。将这些DNA片段和3种酶的混合溶液孵育1h即可。5’外切酶,从5’端开始对DNA进行消化,产生长的黏性末端,这样便于与另外的同源末端进行配对结合;DNA聚合酶,用于修补由5’外切酶产生的缺口,在Gibson组装中用的是Phusion DNA聚合酶,当然,Taq DNA聚合酶也可以用;DNA连接酶,实现共价连接,形成完整的DNA分子。该方法的优势在于这3种酶都可以在同一个温度下发挥功能,可一步完成组装,组装后的质粒可直接用于转化感受态细胞,无需限制性内切酶,通常该方法可组装不超过6个片段。 2010 年,Gibson等科学家使用Gibson组装和酿酒酵母体内同源重组人工合成了1.08Mb的丝状支原体基因组并转入去核的山羊支原体,新移殖的细胞表现出了丝状支原体的性状。2016年,Hutchison 等人先用 PCR 合成一系列丝状支原体基因组片段,每个DNA片段长1.4 kb,然后每5个分成一组,再利用 Gibson 组装技术将5个一组的DNA片段连接成7 kb的单片段,最后组装成完整的基因组。作者只保留了必需基因,将1.08 Mb的基因组缩短到了含有473个基因的 531 kb。2017 年,Esvelt等人将Gibson组装技术与CRISPR/Cas9 技术结合构建了一个PAM质粒文库,用于评估鉴定来自不同物种的一系列不同的Cas9蛋白的活性。该方法在体外组装技术中应用广泛。
图2 基于5′核酸外切酶、DNA 聚合酶及连接酶的Gibson组装法 通过一步等温体外重组两个相邻的DNA片段(红色和蓝色,共用终端序列为黑色),两个DNA片段共价连接成同一个DNA分子。T5外切酶去除双链DNA分子的5’端核苷酸,产生粘性末端,互补单链DNA退火结合在一起,Phusion DNA聚合酶填充缺口,Taq DNA连接酶密封缺口。T5外切酶是不耐热的,在50℃培养期间失活,所以不会与DNA聚合酶竞争DNA模板。 2.2 Golden Gate组装 Golden Gate组装技术是基于非同源重组的代表性技术,其利用Ⅱ型限制性酶 BsaⅠ在识别位点外部切割的特性,设计特异的突出序列同时组装多个片段,且酶切和酶连可以同时进行,原理如图3所示。首先,扩增目的片段,在两端加上BsaⅠ识别序列,同时在识别序列内侧加上不同的4 nt突出部分,相邻片段衎接处的4 nt反向互补配对,然后将片段分别插入酶切前的中间载体,因此一共可以设计256个突出的末端。利用E.coli富集含有不同目的片段的中间载体,将这些载体与最终的载体(含有2个相邻的BsaⅠ酶切位点)混合,加入BsaⅠ和DNA连接酶,同时进行酶切和酶连组装多达9个DNA片段。 2011年,Cermak等人利用Golden Gate高效组装了TALEN敲除系统中的重复序列,整个过程只需要两步:第一步先将最多10个RVDs按顺序组装,每个RVDs可识别1个核苷酸;第二步再将这些RVDs和TALEN系统的其它功能元件如NLS、AD等组装成最终的敲除质粒。2015年,Lauressergues等人使用Golden Gate构建microRNAs表达质粒,探究了苜蓿、拟南芥中2条 microRNAs的转录调控作用及其它5条microRNAs编码的具有调控作用的短肽对植物主根生长的影响。
图3 基于Bsa I内切酶的Golden Gate组装
图3 Golden Gate组装 2.3 同尾酶BioBrick、iBrick 和 C-Brick BioBrick是合成生物学领域一个经典的基因组装技术,BioBrick的原理如图4所示。同尾酶可以识别不同的DNA序列但是能切出相同黏性末端的一类限制性内切酶,但其切开的末端相互连接后不会再形成原酶切位点。其利用同尾酶Xba Ⅰ和 Spe Ⅰ酶切产生相同的粘性末端,酶连后片段连接处会产生86p的疤痕,Xba Ⅰ和Spe Ⅰ都不再识别连接处的疤痕,这样即可在不损失酶切位点的情况下不断组装DNA片段。2014年,国内学者利用归位内切酶I-Sce Ⅰ和PI-Psp Ⅰ可产生相同粘性末端的特点,模仿BioBrick原理设计出一种名为iBrick的DNA 组装方法。归位内切酶识别的序列比普通的限制性内切酶长,在DNA中出现的频 率极低,因此iBrick相比于BioBrick对DNA片段序列几乎没有限制。但较长的识别序列在酶切酶连组装的同时也产生了更长的疤痕,为了解决这个问题,科学家利用一种应用了CRISPR-Cas系统的DNA组装方法C-Brick。C-Brick利用Ⅴ型CRISPR/Cas 系统蛋白Cpf1核酸内切酶,在PAM位点切割产生5nt的粘性末端。该方法利用sgRNA识别长序列的特点避免了对待组装片段序列的要求,同时可灵活设计sgRNA以识别不同的序列,只产生5 bp的疤痕,完美地规避了 BioBrick 的缺点。
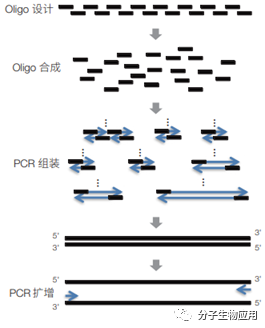
图4 BioBrick技术的组装原理 2.4 基于DNA聚合酶的策略。 该方法首先合成有互补配对重叠区的Oligo,利用PCR扩增的原理,可以对有互补重叠区的Oligo进行延伸连接获得小DNA片段,然后小DNA片段以重叠PCR的方式进行逐步连接获得目的DNA片段,以其作为模板,进一步PCR扩增可以大量获得目的片段(图5)。此方法(polymerase cycling assembly,PCA)无须依赖额外的DNA连接酶,直接从人工合成的Oligo开始组装,操作简单、快速。Stemmer 等人曾用此法实现基因及质粒的一步拼装。Smith等人则 通过增加 Taq 连接酶的步骤用此法合成了 ФX174 噬菌体 的基因组。
图5 基于DNA聚合酶合成法 参考文献: 1.Daniel G Gibson,Enzymatic assembly of DNA molecules up to several hundred kilobases; 2.卢俊南,DNA的合成、组装及转移技术; 3. Liang J,Twin-primer non-enzymatic DNA assembly: an efficient and accurate multi-part DNA assembly method. 4. Li MZ,Harnessing homologous recombination in vitro to generate recombinant DNA via SLIC. 5. Torella JP,Rapid construction of insulated genetic circuits via synthetic sequence-guided isothermal assembly. 6. Torella JP,Rapid construction of insulated genetic circuits via synthetic sequence-guided isothermal assembly. |


 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号