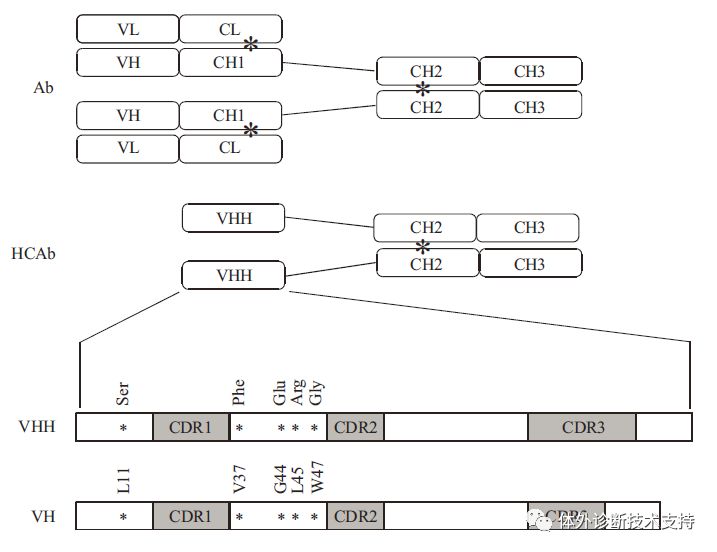
免疫球蛋白(immunoglobulinG,IgG)是由浆细胞分泌的,能对细菌、病毒等抗原进行特异性识别和结合。随着单克隆抗体技术、分子工程和噬菌体展示技术的持续发展,单克隆抗体在免疫试剂、分子显像等疾病诊断方面具有广泛应用前景,并且单克隆抗体药物也已用于心血管病、自身免疫性疾病以及恶性肿瘤等疾病的临床治疗。如,抗癌药物赫赛汀是一种针对乳腺癌HER2 的曲妥珠单克隆抗体,通过阻止人体表皮生长因子(EGF)与HER2的结合,从而抑制癌细胞生长。 Kohler和Milstein在1975 年创建了单克隆抗体技术:通过免疫哺乳动物产生能分泌特异性抗体的浆细胞,与骨髓瘤细胞融合,筛选得到所需细胞株,进而获得单克隆抗体。传统抗体制备过程中不仅需要大量免疫哺乳动物,且需要经过高亲和/特异性分离等繁琐步骤,导至单克隆抗体生产成本高、耗时长,不利于大规模生产。通过基因工程技术在细菌中表达抗原绑定区域(Fab)和单链抗体可变区(scFv),以降低生产成本、提高生产效率。但这些方法生产的Fab 和scFv 抗体普遍存在溶解度低以及片段无法正确折叠/重组等问题,无法满足大规模工业生产的要求。 20 世纪90 年代,Hamers Casterman 等在骆驼科动物的血清中发现了一种缺乏轻链的特殊抗体,称之为单链抗体(HCAb),该结构后来在鲨鱼、蝠鲼和鳐鱼等软骨鱼类的体内也有发现。HCAb 不仅完整保留了抗原结合区域结构,而且保持了与抗原绑定的高亲和力。同软骨鱼类相比,骆驼抗体恒定区与人类的较为相似,容易进行动物免疫实验,因此,目前纳米抗体技术研究主要集中在骆驼科单链抗体及其派生物。 1 纳米抗体结构 传统的IgG 由2 条重链和2 条轻链构成,2 条重链之间以及重链和轻链之间通过二硫键进行连接(图1)。轻链和重链都由可变区和恒定区结构组成,重链的恒定区为CH1、CH2、CH3,可变区为VH,轻链的恒定区为CL,可变区为VL。可变区中的部分氨基酸容易发生突变,称为高变区,由其组成的抗原结合片段位于抗体两臂的末端,决定了IgG的抗原特异性。与传统IgG 相比,在骆驼中发现的单链抗体(HCAb)缺少轻链和重链CH1,只含有重链CH2、CH3 和可变区。HCAb 的抗原绑定部分仅由重链可变区构成,称为纳米抗体(VHH 或Nb),尺寸大约是传统抗体(Ab) 的1/10 (Nbs: 15 kD;Abs:150 kD)。VHH 主要依靠3 个区域与抗原分子结合,称为互补决定区(CDR1、CDR2、CDR3),也称之为高变区(HV1、HV2、HV3),其氨基酸序列具有很强的突变性。高变区之间区域的氨基酸序列变化较小,称为骨架区(FR1、FR2、FR3、FR4),骆驼VHH 骨架区序列与人VH 相比同源性超过80%。
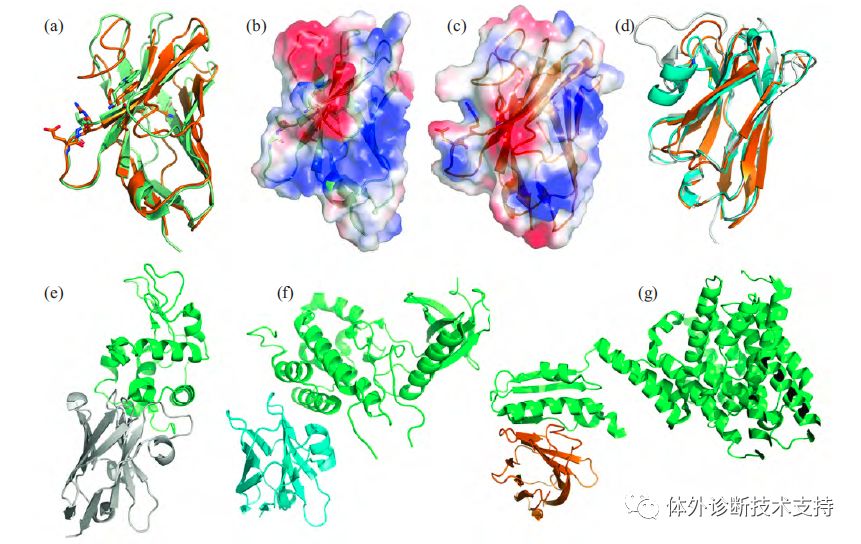
图1 传统抗体、单链抗体及其重链可变区简略结构对比示意图 1.1VH 和VHH 的结构差异 1.1.1骨架区保守序列差异 在传统IgG 重链可变区结构中的Gly44、Leu45和Trp47 这3 个位点位于VH 和VL 的交界处,形成疏水界面,有助于VH 和VL 连接,但VHH 缺少相应的VL 导至上述位点暴露在溶剂中,1994年,Davies 和Riechmann[实验结果显示如果没有VL 的存在,溶液中VH 蛋白质浓度超过1 g/L 时会发生聚集沉淀。而当骆驼源化该区域(Gly44Glu-Leu45Arg(或Leu45Cys)-Trp47Gly)后蛋白质聚集程度显著降低,说明这些位点处于蛋白质表面,改为亲水性残基可增加VH 溶解度。此外,在单峰驼VHH-FR2 区域中Leu11 被替代成Ser11。传统VH中Leu11 位于抗原绑定表位,会与重链CH1 区间接触,是VH 和CH1 间球窝式结构的一部分,有利于抗体的肘部运动。但VHH 中没有CH1 结构,Leu11 没有存在的必要,替代为亲水性氨基酸Ser11 对于溶解度的提高具有重大意义。 1.1.2互补决定区结构差别 除了上面提到的骨架区保守氨基酸差异,VHH 与VH 在高变区(hypervariable,HV)的显著差异主要体现在残基数量及区域结构的不同。首先,VHH-HV1 的残基数扩大到27~35,增加的残基27~30 构成新的β转角,对VHH 的溶解度提升有一定的帮助。此外,溶解酵素和核糖核酸酶A 的VHH 晶体结构显示HV1 长度的增加不仅会对其环结构产生一定的影响还能提高VHH 绑定抗原的能力。其次,研究结果显示,部分骆驼HV2 区间存在与人、鼠相似的发卡结构,而其他HV2 的氨基酸残基数增加,表现出特殊构象,形成多个β转角。VHH 与VH 最显著、最重要的差异在于HV3。虽然不同长度的VHH-HV3 都有发现,单峰驼中长度为16~18 个氨基酸,频率最高,明显长于人和鼠中VH-HV3 的平均长度。更长的HV3 区域扩大了抗原绑定位点的实际表面,弥补了先天缺失的VL 所能提供的抗原绑定表面,使得VHHCDR部分可进入抗原活性中心识别VH 无法识别的位点。通常蛋白质内部氨基酸多为脂溶性,由此可解释为何骆驼的VHH-HV 中脂溶性氨基酸Ala、Ile、Leu 和Val 多于鼠的VH-HV(图2c 针对碳酸酐酶VHH 的CDR3 表面静电分布显示为中性)。 1.2VHH 的结构功能特性 图2a 为抗SLC26 转运蛋白VHH 与抗钾离子通道蛋白Fab 的VH 部分三级结构比较,图2b 和图2c 分别为2 抗体蛋白表面静电图。由图可见,VHH 和VH 蛋白的三级结构大体相同,都由2组反向平行β折叠片层形成类桶装结构(高度为4~5 nm,直径为2~3 nm),与抗原结合的3 个CDR 区间位于同一侧,靠近氮端。但两者表面静电分布却有明显差别,VH 表面极性残基比较集中(如CDR3 区),在FR2 区间(β3-β4 折叠)的几个保守氨基酸为疏水性,表面静电势显中性;而相对应的VHH 表面静电图显示亲水性残基较分散。由此可以阐明单独表达VH 结构域时蛋白易聚合形成包涵体,而VHH 的溶解度高、稳定性好的原因。VHH 作为最小抗原绑定单位,其分子质量约为常规抗体的1/10,结构简单,因此具备较好的组织穿透能力以到达肿瘤病灶,而过多的VHH 能较快由肾脏清除体外,同时,VHH 具有对人体的免疫原性较弱的优势。
图2 Fab 抗体VH 蛋白和VHH 蛋白三级结构及两者结构比较 图2d 为针对不同抗原的3 种VHH 结构比较,图2e、2f、2g 分别为抗溶菌酶、抗细胞周期蛋白G 相关激酶及抗SLC26 转运蛋白的VHH 及其相对应的抗原构图(其中抗原显示为绿色)。由图2 可见,VHH 结构最大差异在于高变区间的残基数量以及结构不同,例如抗溶菌酶的VHH 通过CDR2、CDR3 都参与抗原绑定且CDR2 环伸入溶菌酶结构空腔(图2e),而抗SLC26 的VHH 与抗原结合方式除了由16 个残基组成的VHH-CDR3 环形成一个大的无定形卷曲部分与抗原表面结合外,还存在CDR2 和部分FR2 区间与抗原平行绑定(图2g)。相较于传统抗体,VHH 高变区间结构更为灵活、构型更多,可识别抗原的不同构型及不同表位,具有更广泛的抗原结合方式。此外,抗细胞周期蛋白G相关激酶的VHH 在结构上除了保留与VH 相似的二硫键(Cys22-Cys93)外,同时在FR2 和CDR3 之间新增了一个二硫键(Cys47-Cys99),并且CDR3区间因增加一个α螺旋而表现出新的环构型(图2f),由此稳定并束缚该纳米抗体的抗原绑定区结构。 2 纳米抗体来源及纳米抗体库构建 单链抗体最先在骆驼科动物体内发现,主要由免疫单峰驼和羊驼等骆驼科动物获得。免疫过程为期5~6 周,第1 周注射与完全佐剂混合的抗原,接下来每周注射1 次等体积混合不完全佐剂的抗原以刺激骆驼产生抗原特异性的、高亲和性的抗体。此外,还可以通过一次使用多种抗原免疫骆驼来减少成本,提高制备效率。最终免疫的效果往往由抗原性质决定,相较于非极性半抗原,体积较大的抗原能够引起更强的免疫反应。 2.1VHH 的制备 2.1.1血浆中提取 单峰驼血清中IgG 的总量约为5~10 g/L,其中IgG1 为传统抗体,IgG2 和IgG3 为单链抗体。IgG1 和IgG3 通过亲和吸附绑定于蛋白G 亲和色谱柱(IgG2 不与蛋白G 亲和柱绑定),而后用150mmol/LNaCl, 0。58% 醋酸(pH 4。5) 的缓冲液淋洗分离出IgG3,第一步中的洗脱液再次通过蛋白G 亲和色谱柱以确保吸附其中剩余的少量IgG1 和IgG3。收集仅含IgG2 的流出液,采用蛋白A 亲和色谱柱分离纯化。纯化后的IgG2 和IgG3 通过蛋白酶(如胃蛋白酶、胰蛋白酶、金黄色葡萄球菌V8 蛋白酶等)酶解其VHH 和CH2 之间的铰链,再用蛋白A凝胶过滤亲和色谱绑定酶解所得的CH部分,而VHH 随着淋洗液流出,达到分离纯化目的,即从血清中纯化得到VHH。 2.1.2VHH 重组及库的筛选 免疫建库法主要包括免疫骆驼、提取VHH 基因、建立特异性纳米抗体库以及运用噬菌体展示技术对库进行筛选4 大步骤(图3)。首先,从已免疫的单峰驼中抽取20~30 ml 外周血,获取效应B 细胞并从中提取总RNA,运用逆转录PCR 技术获取cDNA。由于VHH 片段PCR 扩增过程中可能引入VH 基因,直接影响到纳米抗体库的构建筛选,故需设计合适引物以扩增高纯度VHH 片段。利用抗体不同的同型,如IgG1 与IgG2、IgG3 的铰链存在较大差异的特点,van der Linden 等设计一套引物可扩增出所有种类骆驼的VHH 部分,而Maass 等设计了仅针对羊驼的VHH 区间的引物。另一种VHH 片段扩增方法利用了VHH 与VH基因差异性(图1)以及所有的VHH 基因具有相同的外显子和同源序列的特性。采用巢式PCR 技术,先用一对引物将VHH 至CH2 以及VH 至CH2 之间的片段扩增,得到不含有CH1 或含CH1 两种不同长度的DNA 片段,通过琼脂糖凝胶电泳可以将不含CH1 的HCAb 重链部分分离出来(约600 bp),接下来利用一对含限制性内切酶位点的巢式引物,将针对性扩增VHH 片段。该方法可用于单峰驼、无峰驼、双峰驼以及羊驼IgG3 中VHH 片段的扩增。将PCR 扩增所得VHH 基因转入特定的表达载体中,便得到了包含完整抗原绑定区域的VHH库。相较于传统抗体建库,VHH 库的建立过程更加简单,所需的库容量也更小,约106~107 个克隆(传统抗体至少需要1010 个克隆)。
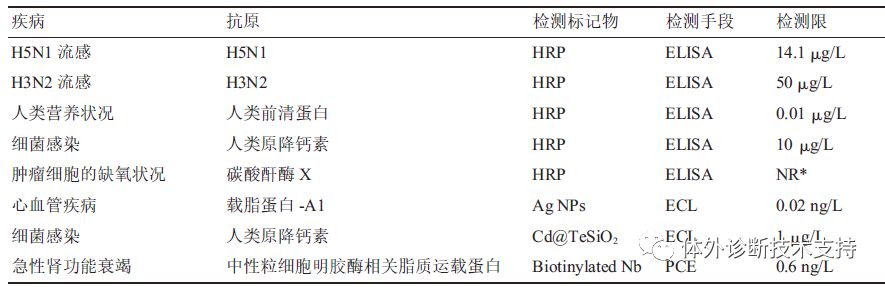
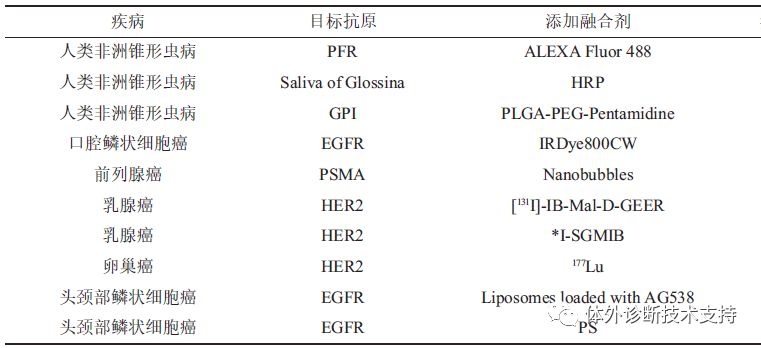
图3 VHH 制备流程 目前VHH 库的筛选方法除了利用酶联免疫法对细胞表达产物进行筛选外,通过选用线性噬菌体(包括M13、f1、Fd、ft 等株型) 进行噬菌体展示以筛选阳性克隆的方法更为常用。该方法首先用辅助噬菌体超感染大肠杆菌VHH 库建立噬菌体展示库,在大肠杆菌中扩大培养后分离得到在衣壳蛋白上展示VHH 蛋白的噬菌体。接下来用固定的抗原对噬菌体进行筛选,洗去未与抗原绑定的噬菌体,而与抗原绑定的噬菌体通过低pH 和高浓度尿素洗脱,中和后重新感染细菌。该筛选操作过程可以进行多次循环(一般为2~3 次)以获得高亲和特异性VHH。通过基因工程将VHH 克隆至表达载体,表达纯化后得到可溶性VHH 蛋白。从免疫的动物中获得血样到最终得到高亲和的抗原特异性VHH 抗体整套操作用时仅3 个月不到,与传统单克隆抗体的制备、筛选过程相比更加简便高效。 2.1.3人工合成VHH 库的构建 对于毒性太强、不具免疫原性、提取量不足而不能产生免疫反应的抗原,是无法通过传统免疫法制备抗体。而利用简并密码子NNK(随机改变库的寡核氨酸)增加VHH 中CDR3 核苷酸多样性,并通过重叠延伸PCR 技术,重组、扩增目的基因,可构建针对上述特殊抗原的VHH 库。 3 纳米抗体技术在疾病诊断与治疗中的应用 利用纳米抗体对抗原的特异性和高亲和性,在疾病诊断中,纳米抗体技术多采用酶联免疫法(enzyme-linked immunosorbent assay, ELISA) 进行检测。在基本的酶联免疫法中,聚苯乙烯和聚氯乙烯常作为固相载体用于抗原的固相化,该类载体对蛋白质有较强的吸附能力,并能保留抗原原有的免疫学活性。由于抗原通过疏水基团间非特异性的物理吸附被固定在载体上,为避免干扰物质的再吸附,常用高浓度牛血清蛋白和脱脂奶粉作为封闭溶液填充载体空隙,并用洗涤液洗涤去除杂质和未结合蛋白。选用高效灵敏的酶(如:HRP 和葡萄糖氧化酶)和高纯度的抗体制备酶标抗体,不仅保留了抗体的免疫活性,同时具备酶的高效催化能力。加入酶标抗体进行反应稳定后,用洗涤液洗涤,最后加入酶底物进行检测。常用的HRP 显色底物是以四甲基联苯胺(TMB)为氢供体、双氧水为氢受体,二者按一定比例混合后作为TMB 工作液,用硫酸终止酶催化后,底物从蓝色变成黄色,可在最适吸收波长450 nm 处定量分析。 除了上述提及的酶联免疫基本构建方法,实际使用中较为广泛的是双抗体夹心法,对应纳米抗体的“三明治”结构(即NbⅠ- 抗原-NbⅡ形式),此结构包含了两种能和抗原不同表位结合的纳米抗体。纳米抗体“三明治”结构已用于多种疾病的诊断(表1)。将NbⅠ固定于载体上(用于捕获待测抗原)的方式存在以下两种:a。磁珠纳米颗粒键合,即将金属氧化物的纳米颗粒和磁性颗粒结合,利用共价键键合绑定NbⅠ;b。生物酰化处理,即融合表达NbⅠ与生物酰素,利用生物酰素与链霉亲和素的结合能力间接绑定NbⅠ。 针对NbⅠ捕获的抗原,通过加入NbⅡ用于间接检测抗原的浓度。其中对于NbⅡ浓度的检测方法多样,较为常见和简易的方法为酶标法。除此之外,可利用具有较高灵敏度的纳米金属或是金属氧化物与NbⅡ的绑定,检测NbⅡ上标记物反应前后的电化学性质变化进而计算出NbⅡ的浓度。Li报道了还可采用光敏平板,配合金属氧化物修饰的NbⅡ,检测反应前后在光照下呈现出的不同电信号,依次检测即可计算出相应的NbⅡ浓度。 3.1 纳米抗体在肿瘤诊断中的应用 表皮生长因子接受子(epidermalgrowth factor receptor,EGFR)是一种跨膜糖蛋白,可促进细胞增殖,抑制细胞的程序性凋亡,使癌细胞具攻击性,更容易发生转移,过量表达于乳腺癌、脑癌等实体肿瘤中。研究表明,作为EGFR 家族代表之一的HER2 在20%~30%的乳腺癌、4%~6%的非小细胞肺癌、20%~24%的胃癌以及结肠癌和卵巢癌中都会过度表达。因此EGFR 家族蛋白常作为抗癌研究的靶标热点。目前,临床上采用以放射性元素或其他方法标记的单克隆抗体作为显像剂的分子显像技术确定病人体内肿瘤分布情况。但单克隆抗体在血液中的保留时间过长,导至检测背景值过高,显像效果不理想。而针对肿瘤的特异性纳米抗体可以更好地渗入癌变组织,识别传统抗体无法靶向的活性位点,是理想的显像剂载体。 近红外(near-infrared, NIR)波长范围(650~800 nm)内的光被人体组织吸收的较少,穿透性较好,且其非特异性背景信号较低,故可作为癌症检测的重要显像手段。 van Driel 等使用NIR 荧光-FLARE显像系统进行小鼠体内口腔鳞状细胞癌(oral squamous cell carcinoma,OSCC)的显像研究。耦合了荧光团IRDye800-CW的纳米抗体可以特异性靶向并绑定OSCC 中过量表达的EGFR,能清晰地显示OSCC 及颈部淋巴结的转移情况,在临床上有着十分可观的应用前景。 超声分子成像技术采用特异性抗体标记的显像分子探针,通过超声检测显像剂在目标部位的聚集情况,获得分子或活细胞层面上的影像。肿瘤中新生血管的最大孔径为380~700 nm,而广泛使用的超声显像剂直径为1~10 μm,难以到达肿瘤部位。Fan 等[36]研究结果显示,通过将生物素标记的抗前列腺特异性膜(prostate-specific membrane antigen,PSMA)的纳米抗体与超声纳米泡耦合方法,可构建能高效绑定小鼠体内前列腺癌细胞的超声显像分子(平均直径为(487。60 ± 33。55) nm)。 赫赛汀(曲妥珠单抗)是治疗乳腺癌的首个分子靶向药物,主要针对HER2 阳性的癌症病人,但对HER2 表达较少或表达多样化的病人治疗效果较差,且其价格昂贵,长期使用会使患者产生耐药性,用曲妥珠单抗治疗前需准确测定患者体内HER2 表达情况。Pruszynski研究表明,放射性碘化试剂SGMIB 标记针对HER2 的Nb(5F7GGC)为显像剂(5F7GGC-125/131I-SGMIB)注入异种移植小鼠,肿瘤吸收2 h 达到峰值,且肿瘤与正常组织吸收比值可达到50∶1,能准确反应癌症中HER2 表达情况。此外,由于纳米抗体能更快地被正常组织清除,可与半衰期较短的放射性核素68Ga、18F 或211At 结合,用于分子显像或靶向治疗。
3.2 纳米抗体在肿瘤治疗中的应用 化学药物疗法是临床上常用的肿瘤治疗方法之一, 烷化剂、抗代谢药物、抗癌抗生素等化疗药物在体内杀伤肿瘤细胞的同时,也会对正常细胞产生非特异性毒性。用于治疗结肠癌和直肠癌的第三代铂类抗癌药物奥沙利铂在体内有50%与红细胞结合,从而造成一定的周围神经毒性。相较于“裸药”,纳米抗体与高分子液泡/脂质体耦合的药物载体可以提高药物溶解度、稳定性;利用纳米抗体靶向传递药物;通过改变高分子液泡/脂质体的双亲结构达到控制药物释放的目的。der Meel 等报道了将类胰岛素生长因子1 接受子(insulin-like growth factor 1 receptor,IGF-1R)抑制剂(AG538)封闭在脂质体内,并将其与抗EGFR的Nb(EGa1)耦合形成EGa1-AG538-liposomes 结合物。研究结果显示,该药物载体可达到同时阻遏EGFR 和活化IGF-1R 的双重效果,有效抑制肿瘤细胞扩散,是治疗由EGFR 和IGF-1R 过度表达引起肿瘤的理想药物。 放射性免疫治疗(RIT)在癌症治疗中应用广泛,当采用传统单克隆抗体mAb 作为放射性元素载体时,因其大分子量造成药物在肿瘤区间分布、吸收缓慢并对肿瘤的渗透性不理想,而降低了RIT 效率。D'Huyvetter报道177Lu 标记的抗HER2 的纳米抗体可有效抑制小鼠体内过量表达HER2 肿瘤的增长,且不会非特异性辐射健康组织,同时组织病理学数据显示其对肾脏无毒性,证明该放射源标记的纳米抗体可用于放射性靶向治疗。 光动力疗法(PDT)因其创伤小、选择性强、适用性范围广等优点在肿瘤治疗中有良好应用。PDT是指将光敏剂(PS)静脉注射到病人体内,经光照活化后的PS 使体内O2形成活性氧物质,穿透肿瘤细胞线粒体膜或核膜,造成DNA 和蛋白质的损伤。但是疏水性的PS 分子在体内容易聚集,并具有脱靶毒性,从而破坏正常细胞。Heukers通过筛选纯化出高特异性针对EGFR 的Nb,并以此构建Nb-PS 偶联物,使PS 能更快更均匀地分布在肿瘤内,从而能快速高效地靶向杀死肿瘤细胞,在肿瘤治疗中有十分广阔的应用前景。 Nb 不仅是抗癌药物载体的重要组成部分,也可直接作为药物治疗肿瘤。Farajpour通过噬菌体建库、筛选、纯化得到针对人体血管内皮生长因子(VEGF)的Nb。体外细胞实验显示,不同于传统单抗以空间位阻的方式阻碍VEGF 与接受子结合的方式,Nb 可靶向单抗无法识别的活性位点,即VEGF 与接受子的结合位点,进而抑制VEGF 引导的内皮细胞增生。 3.3 纳米抗体在非洲锥形虫病诊断治疗中的应用 人类非洲锥形虫病(humanAfrican trypanosomiasis,HAT)是一种通过采采蝇的叮咬传播,由布氏锥虫引起的慢性感染病,导至36 个非洲国家超过6000万人口受到威胁。现在该病的诊断主要是采用间接酶联法或间接荧光法检测血清中的抗体,这种方式无法区分被测对象处于新近感染或已治愈中的哪种情形。相比之下,以抗原为对象进行检测准确度更高。然而,锥形虫拥有庞大的编码可变表面糖蛋白(variant surface glycoprotein,VSG)的基因组,可激活不同VSG基因产生不断变化的糖蛋白以躲避宿主免疫系统,增加了以VSG 作为抗原检测锥虫病的难度。 Obishakin的研究表明,非洲锥虫病的致病原锥形虫的鞭毛杆蛋白(paraflagellar rod protein,PFR)可以成为诊断该病的有效抗原。实验采用T。 Evansi STIB 以及4 种不同T。 Evansi 伊式锥虫混合裂解液免疫骆驼,并构建特异性Nb 库。经纯化、筛选得到的Nb392 用荧光染料ALEXA 标记,通过免疫荧光检测方法以及RNAi 技术确认Nb392可以特异性结合鞭毛蛋白PFR1。Caljon 等用采采蝇唾腺膜蛋白膜提取物免疫骆驼,构建特异性纳米抗体库。经筛选得到的TsalNb5 和TsalNb11 与辣根过氧化物酶(HRP)耦合,用于竞争性免疫分析。结果显示TsalNb5 以及TsalNb11 能够特异性识别采采蝇,而不与厩螯蝇、马蝇等其他品种发生交叉反应。上述方法不仅可对采采蝇唾液蛋白进行微量检测,也可作为非洲锥虫病病原体通过载体传播过程的风险指标,有利于对该病传播途径的干预和控制。 纳米抗体除用于非洲锥虫病的检测外,也可作为药物载体靶向特定位点从而在该病的治疗中发挥重要作用。由于锥形虫具有上述不断改变VSG 的特性,针对该病的疫苗难以制成,故控制非洲锥虫病只能通过化学治疗得以实现。但是,用以治疗该病的药物种类少且副作用大,仅有5 种药物在临床上被使用,戊烷脒(pentamidine)是其中之一。其主要用于冈比亚布氏锥虫引起的HAT 第一阶段的治疗。Arias 等构建了Nb-pentamidine-PLGA 药物载体复合物,该纳米颗粒利用NbAn33 靶向锥形虫表面相对保守的糖磷脂酰肌醇(GPI)免疫表位,以及聚乳酸- 羟基乙酸共聚物(PLGA)包裹戊烷脒,通过锥形虫表面鞭毛口袋的胞吞作用摄入药物从而起到治疗作用,大大增加了戊烷脒的药效。
4 纳米抗体技术应用展望 从1993 年报道首个在骆驼体内天然存在的单链抗体的发现以来,重链可变区(又称纳米抗体)由于具有分子质量小、结构稳定、溶解度高、对抗原的高亲和力以及能在微生物中高量表达等特性,结合噬菌体展示技术,日渐成熟的纳米抗体技术已被广泛应用于酶联免疫、生物传感器检测等研究领域。目前纳米抗体技术主要用于疾病的体外诊断,但已有纳米抗体药物处在临床二期实验的相关报道,如Ablynx 公司研制的ALX-0061 通过与IL-6(促炎细胞因子)特异性结合从而完全抑制由IL-6 引起的风湿性关节炎炎症。随着纳米抗体在分子显像剂、药物载体等方面的发展,我们有理由相信纳米抗体技术将会在人类癌症等疾病的临床诊断和治疗中发挥越来越重要的作用。此外,纳米抗体还可作为研究工具用于:a.与固相载体相连用于捕获目标蛋白辅助蛋白质的分离纯化;b.以共结晶方式获得常规方法无法得到的难溶蛋白质晶体;c。进入细胞核内与转录因子结合从而驱动DNA的转录。纳米抗体可以构建成双价(针对同一抗原)或双特异性(针对不同抗原)抗体,这将提供一种即插即用的检测手段,有利于纳米抗体技术广泛地应用于疾病的临床快速诊断。 参考: 1. 郭海涛. 单链抗体的研究进展及其应用.2015 3. 周陶然.纳米抗体技术及其在疾病诊断和治疗中的应用.2016 4. L. Eyer, K. Hruska.Single-domainantibody fragments derived from heavy-chain antibodies: a review.2012 |

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号