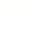用户名
UID
Email
密码
记住
立即注册
找回密码
只需一步,快速开始
微信扫一扫,快速登录
开启辅助访问
快捷导航
门户
Portal
社区
BBS
资讯
会议
市场
产品
问答
数据
专题
帮助
签到
每日签到
企业联盟
人才基地
独立实验室
产业园区
投资机构
检验科
招标动态
供给发布
同行交流
悬赏任务
共享资源
VIP资源
百科词条
互动话题
导读
动态
广播
淘贴
法规政策
市场营销
创业投资
会议信息
企业新闻
新品介绍
体系交流
注册交流
临床交流
同行交流
技术杂谈
检验杂谈
今日桔说
共享资源
VIP专区
企业联盟
投资机构
产业园区
业务合作
投稿通道
升级会员
联系我们
搜索
搜索
本版
文章
帖子
用户
小桔灯网
»
社区
›
C、IVD技术区
›
原料技术
›
如何理解酶结构和酶活性之间的关系?
图文播报
2026庆【网站十三周
2025庆【网站十二周
2024庆中秋、迎国庆
2024庆【网站十一周
2023庆【网站十周年
2022庆【网站九周年
返回列表
查看:
13419
|
回复:
4
[分享]
如何理解酶结构和酶活性之间的关系?
[复制链接]
风云
风云
当前离线
金桔
金币
威望
贡献
回帖
0
精华
在线时间
小时
发表于 2024-9-14 10:02
|
显示全部楼层
|
阅读模式
登陆有奖并可浏览互动!
您需要
登录
才可以下载或查看,没有账号?
立即注册
×
急急急拜托了各位
原文地址:https://www.zhihu.com/question/590007274
回复
举报
卡卡
卡卡
当前离线
金桔
金币
威望
贡献
回帖
0
精华
在线时间
小时
发表于 2024-9-14 10:03
|
显示全部楼层
正确理解酶的专一性及实验
正确理解酶的专一性
酶专一性是相对的,而且教材上的酶的专一性仅仅是一个不完全归纳,还不能证明酶的专一性,这也是科学研究结论得出所注意的,严谨地说,假设成立需要用完全归纳法。这也诠释了教材为什么用“探究酶的专一性”道理。
问题1:探究“酶的专一性”还是证明“酶的专一性”?
教材通过“淀粉酶对淀粉和蔗糖的水解作用”实验,让学生体会到:淀粉酶能够催化淀粉的水解,而不能催化蔗糖的水解。这有助于学生理解酶的专一性。但是,这个实验并不能证明酶具有专一性。
因为该实验能够得出这样的结论:淀粉酶能催化淀粉的分解,而不能催化蔗糖的水解。此处实验结果提出属于
不完全归纳
,即淀粉酶具有专一性,它只能催化淀粉的分解,而不能催化其它物质的分解。要想证明这个“结论”,我们需要测试蔗糖以外的其它物质,如脂肪、蛋白质、核酸以及其它糖类等物质,结果发现对于自然界的所有物质,淀粉酶都只能催化淀粉的分解,对其它物质不起作用。
这样我们才能说淀粉酶的专一性得到了证明。完全归纳的结果才是真实可靠的,只有完全归纳才能做到证明。
教师在讲授实验时,一定要注意我们的认识通常都是基于不完全归纳,要慎用“证明”,多用“验证”。像淀粉酶对淀粉和蔗糖的水解作用这个实验,我们可以说这个实验验证了淀粉酶具有专一性,而不说证明了淀粉酶。
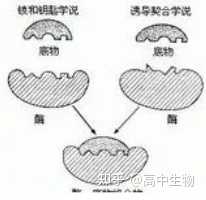
问题2:酶的专一性是相对的。
酶的专一性表现在两大方面:
1.立体异构专一性。一种酶只能对一种立体异构体起催化作用。
2.结构专一性。
(1)绝对专一性:有些酶作用底物只有一个,而不作用于任何其它物质。这种专一性我们称为“绝对专一性”。
(2)相对专一性:有些酶对底物的要求比绝对专一性低,作用对象不只是一种底物,这种专一性又称为“相对专一性”。
相对专一性可分为基团专一性和键专一性。
DNA连接酶的专一性应该属于“键专一性”,在两个断开的DNA片断之间形成磷酸二酯键。
并非所有的酶分子都具有高度专一性。例如,在食品工业中使用的某些蛋白酶虽然选择性地作用于蛋白质,然而对于被水解的肽键都显示相对较低的专一性。当然,也有一些蛋白酶显示较高的专一性,例如胰凝乳蛋白酶优先选择水解含有芳香族氨基酸残基的肽键。
专一性催化机理
在研究酶促反应的机理时,不得不提到过渡态理论或中间产物理论。1913年生物化学家Michaelic和Menten提出了酶中间产物理论。
他们认为:酶降低活化能的原因是酶参加了反应而形成了酶-底物复合物。这个中间产物不但容易生成,而且容易分解出产物,释放出原来的酶,这样就把原来能阈较高的一步反应变成了能阈较低的两步反应。由于活化能降低,所以活化分子大大增加,反应速度因此迅速提高。
这个理论的关键是认为酶参与了底物的反应,生成了不稳定的中间主产物,因而使反应沿着活化能较低的途径迅速进行。事实上,中间产物理论已经被许多实验所证实,中间产物确实存在。
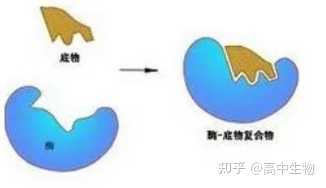
(1) 锁和钥匙学说1984年Emil Fischer提出锁和钥匙模型。
该模型认为,底物的形状和酶的活性部位被认为是彼此相适合,像钥匙插入锁孔中,认为两种形状是刚性的和固定的,当正确组合在一起时,正好互相补充。
(2) 诱导契合学说
后来许多化学家发现,许多酶的催化反应并不符合经典的锁和钥匙模型。1958年Daniel E. Koshland Jr. 提出了诱导契合模型,底物的结合在酶的活性部位诱导出构象的变化。
该模型的要点是:当底物与酶的活性部位结合,酶蛋白的几何形状有相当大的改变;催化基团的精确定向对于底物转变成产物是必需的;底物诱导酶蛋白几何形状的改变使得催化基团能精确地定向结合到酶的活性部位上去。
回复
支持
反对
举报
检验医师
检验医师
当前离线
金桔
金币
威望
贡献
回帖
0
精华
在线时间
小时
发表于 2024-9-14 10:04
|
显示全部楼层
在我们体内,一直有一只神奇的“军队”默默工作,它们便是酶。虽然看似微小无形,但却扮演着举足轻重的角色。
酶的组成,大多数是蛋白质,少数是核酸,或是说酶是一种有机物。
下面小编带你解开酶的神秘面纱,探索它独特的特性。
01
酶是一种催化剂,
能够加速化学反应的速度而不被消耗
1个酶分子在1min内,能引起数百万个底物分子转化为产物,酶的催化能力比一般催化剂的催化能力大10的7次方~10的13次方倍。
酶催化反应的另一个特点,就是酶对底物高度的专一性。一种酶只能催化一种或一类物质反应,即酶是一种仅能促进特定化合物、特定化学键、特定化学变化的催化剂。
02
数以千计的酶存在于人体内,
这便是酶的多样性。
每一种都有其专属任务。
例如:消化酶帮我们分解食物,使其能够被身体吸收和利用;代谢酶参与能量产出和排出的过程;抗体酶会加强免疫系统的功能等。正是这些酶的协同作用,使我们身体能够正常运转。
酶是生物催化剂,它很神奇,神就神在它高度专一,灵敏异常,它效率惊人,使普通催化剂望尘莫及。然而酶又非常娇气,高温、强酸、强碱、重金属盐都是它的克星,因为它毕竟是蛋白质。
03
酶的另一特性,则是高度专一性
举个生活中很常见的例子来形容,就好比“一把钥匙只能开一把锁”。
如果把一个小小的细菌细胞放大10亿亿倍,也不过一个集装箱那么大。
把它打开,就可以看到里面同时进行着1500-2000种化学反应,1000多种酶在忙碌不停地对这些反应进行催化。每一种酶只负责一种反应,自己找准对象,与之结合在一起,绝不会认错。有多少种反应就有多少种酶在尽责,不多也不少。
其实,酶在生命活动中的高度专一性,可以归结为两个方面:
◆ 一方面,酶忠诚于自己的作用对象,绝不错认。
◆ 另一方面,酶忠诚于自己的催化职责——促进特定化学反应顺利进行,并且完成。
04
酶究竟在什么温度范围,
表现出最大的活力呢?
动物细胞一般在37—50℃之间,植物细胞在50一60℃之间最适合。酶在常温常压下就发生催化作用的本领,为人类提供服务的同时,也会给人类带来麻烦。
比如,喝生水容易闹肚子,是因为生水中的细菌,在酶的参与下,细菌迅速繁殖,如大肠杆菌每20分钟就产生一代。大量细菌在人的胃肠里生儿育女,兴风作浪,当然就容易得病。
喝开水就没事。因为开水达到100℃时,细菌体内的蛋白质(包括酶)都凝固起来了,这样的水喝下去就平安无事。刚买回的鱼肉、蔬菜等要放进冰箱保鲜,是为了降低常温,抑制细菌体内的生化反应。因为温度为0℃时,酶的催化反应速度很慢。
医院对衣被、餐具、器械的高温消毒,又是利用了酶、蛋白质的热变性特点来杀死细菌。降温也好,升温也好,都是为了消除酶在常温催化给我们制造的麻烦。
看到这,相信大家对于酶,已经有了充分的认知。总结一下,酶作为生物体内的催化剂,在我们体内发挥着重要的作用,参与各种生化过程,维持身体的正常功能。
而酶的神奇力量远不止于此,科学家们正不断探索酶的新功能和应用,为人类的健康和发展带来无限的希望。
回复
支持
反对
举报
同花顺
同花顺
当前离线
金桔
金币
威望
贡献
回帖
0
精华
在线时间
小时
发表于 2024-9-14 10:05
|
显示全部楼层
(一) 酶分子的结构特点
A 结合部位 Binding site
•
酶分子中与底物结合的部位或区域一般称为结合部位。
B 催化部位 catalytic site
•
酶分子中促使底物发生化学变化的部位称为催化部位
。
•
通常将酶的结合部位和催化部位总称为酶的活性部位或活性中心。
•
结合部位决定酶的专一性,
•
催化部位决定酶所催化反应的性质。
酶分子上具有一定空间构象的部位,是酶分子中直接与底物结合,并和酶催化作用直接有关的部位,称为酶的活性中心。
酶的活性中心的特点
1、活性中心仅占据整个酶体积的很小一部分;
2、是一个具有三维构型的实体;
3、底物和酶的活性中心的结合力很弱;
4、诱导适应过程;
5、含有催化基团和结合基团。
必需基团
Ø
活性中心内的必需基团
结合基团(binding
group)
与底物相结合;
催化基团(catalytic
group)
催化底物转变成产物
Ø
活性中心外的必需基团
维持酶活性中心应有的空间构象所必需。
酶活性中心的必需基团
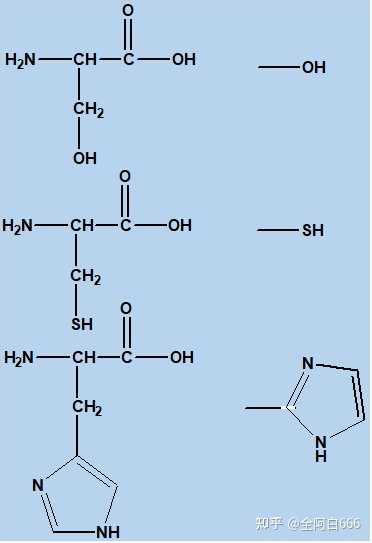
•
主要包括:
亲核性基团
:丝氨酸的
羟基
,半胱氨酸的
巯基
和组氨酸的
咪唑基
。
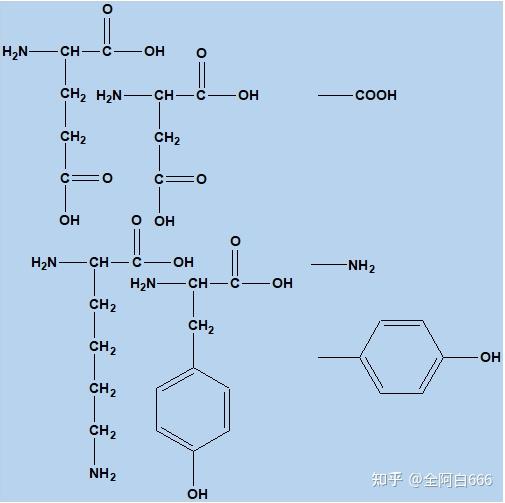
•
酸碱性基团:门冬氨酸和谷氨酸的羧基,赖氨酸的氨基,酪氨酸的酚羟基,组氨酸的咪唑基和半胱氨酸的巯基等。
活性部位的基团都是必须基团,但是必需基团还包括那些在活性部位以外的,对维持空间构象必须的基团!
C 别构部位 Regulatory site
•
酶分子中存在着一些可以与其他分子发生某种程度的结合的部位,从而引起酶分子空间构象的变化,对酶起激活或抑制作用。
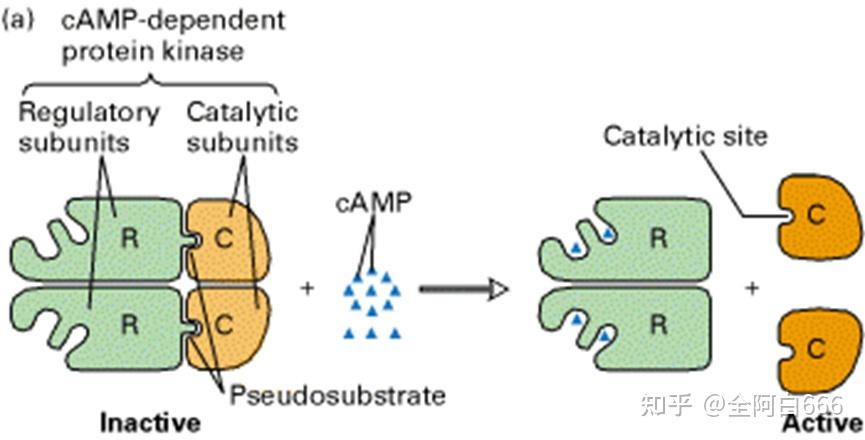
(二) 酶原的激活
•
处于无活性状态的酶的前身物质
就称为酶原。
•
酶原在一定条件下转化为有活性的酶的过程称为
酶原的激活
。这个过程实际上就是酶的活性部位形成或暴露的过程。
•
酶原的激活过程通常伴有酶蛋白一级结构的改变。
•
酶原激活的机制为:酶原分子一级结构的改变导致了酶原分子空间结构的改变,使催化活性中心得以形成,故使其从无活性的酶原形式转变为有活性的酶。
•
酶原激活的生理意义在于:保护自身组织细胞不被酶水解消化。
回复
支持
反对
举报
继续前进
继续前进
当前离线
金桔
金币
威望
贡献
回帖
0
精华
在线时间
小时
发表于 2024-9-14 10:06
|
显示全部楼层
谢谢邀请,虽然这不是我专业所长,但我尽力回答。如有错误,欢迎指出。
了解酶的结构与活性之间的关系,是充分认识酶在生化反应中的作用的必要条件。酶是一种蛋白质,它作为催化剂加速生物体内的化学反应而不被消耗或改变。酶的结构与其活性和功能有着错综复杂的联系。这里有一些关键点来帮助你理解这种关系:
一级结构
:酶的一级结构是组成蛋白质的氨基酸的线性序列。这个序列决定了酶的整体折叠模式,并最终决定了它的三维结构。氨基酸序列是至关重要的,因为即使是微小的变化也会改变酶的活性或使其失去功能。
二级结构
:二级结构是指蛋白质内部的局部折叠模式,如α -螺旋和β -薄片。这些结构是由多肽链中氨基酸之间的氢键稳定的。二级结构有助于酶的整体形状和稳定性,这反过来又影响其催化活性。
三级结构
:三级结构是蛋白质的整体三维形状,由二级结构元素之间的相互作用产生。这种结构水平对酶的功能至关重要,因为它形成了酶的活性位点,底物在此结合并发生化学反应。活性位点的形状和化学性质使酶能够特异性地识别并与底物结合,确保只发生正确的反应。
第四结构
:一些酶由多个多肽链(亚基)组成,它们组装成一个功能酶。四元结构指的是这些亚基之间相对的排列。亚基之间的相互作用可以通过影响酶的整体形状、稳定性和活性位点的可及性来影响酶的活性。
酶-底物特异性
:酶的结构决定了它的特异性,即识别和结合特定底物的能力。活性位点的形状和化学性质与底物互补,允许酶选择性地结合和催化反应。这种酶-底物相互作用的“锁和钥匙”或“诱导匹配”模型确保酶只催化特定的反应。
酶辅因子和假体基团
:一些酶需要额外的非蛋白质成分,如金属离子或有机分子,才能完全发挥功能。这些辅因子和假体基团对于稳定酶的结构或直接参与催化反应是必不可少的。
变构调节
:变构酶有额外的调节位点,与活性位点分离,其他分子可以结合在那里调节酶的活性。这些分子被称为变构效应物,可以通过诱导其结构中的构象变化来激活或抑制酶,从而影响活性位点的形状或可达性。
酶结构和酶活性之间的关系就像一把锁和钥匙的关系。酶的结构就像一把锁,而酶活性就像是开锁所需的钥匙。只有当钥匙的形状和锁的形状完全匹配时,锁才能被顺利地打开。
类比这个例子,酶的结构也有一个所谓的“活性位点”,就像锁的插孔,只有与其相互匹配的底物才能够在此处催化反应。底物的形状和酶活性位点的形状应该彼此适配,就像钥匙和锁的关系一样。如果底物的形状与酶活性位点的形状不相符,则底物无法与酶结合,反应也无法发生。
此外,酶的结构和酶活性之间的关系还可以用另一种比喻来解释。酶的结构就像一个3D拼图,而酶活性就像是唯一正确的拼图方案。只有当每一块拼图正确地放在了它应该放置的位置时,整个拼图才能被完美地组装起来。
类比这个例子,酶的氨基酸序列就像是拼图的各个拼块,而酶的二级和三级结构则是各个拼块之间的相互作用。只有当氨基酸序列和二级/三级结构正确地组合在一起,才能形成一个正确的酶结构,才能让酶完全发挥其催化作用。
总之,酶的结构和酶活性之间的关系可以用“锁和钥匙”或“3D拼图”的比喻来形象地解释。只有当酶结构和酶活性相互适配时,酶才能够完美地催化反应。
回复
支持
反对
举报
返回列表
发表回复
高级模式
B
Color
Image
Link
Quote
Code
Smilies
您需要登录后才可以回帖
登录
|
立即注册
本版积分规则
发表回复
回帖后跳转到最后一页
浏览过的版块
科技前沿
学术培训
关闭
官方推荐
/3
AI助手<小桔子>来了!
欢迎来交流,可以回答IVD行业各类问题!
查看 »
IVD业界薪资调查(月薪/税前)
长期活动,投票后可见结果!看看咱们这个行业个人的前景如何。请热爱行业的桔友们积极参与!
查看 »
小桔灯网视频号开通了!
扫描二维码,关注视频号!
查看 »
返回顶部
快速回复
返回列表
客服中心
搜索
洽谈合作
关注微信
微信扫一扫关注本站公众号
个人中心
个人中心
登录或注册
业务合作
-
投稿通道
-
友链申请
-
手机版
-
联系我们
-
免责声明
-
返回首页
Copyright © 2008-2024
小桔灯网
(https://www.iivd.net) 版权所有 All Rights Reserved.
免责声明: 本网不承担任何由内容提供商提供的信息所引起的争议和法律责任。
Powered by
Discuz!
X5.0 技术支持:
宇翼科技
浙ICP备18026348号-2
浙公网安备33010802005999号
快速回复
返回顶部
返回列表

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 






 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 发表于 2024-9-14 10:02
发表于 2024-9-14 10:02