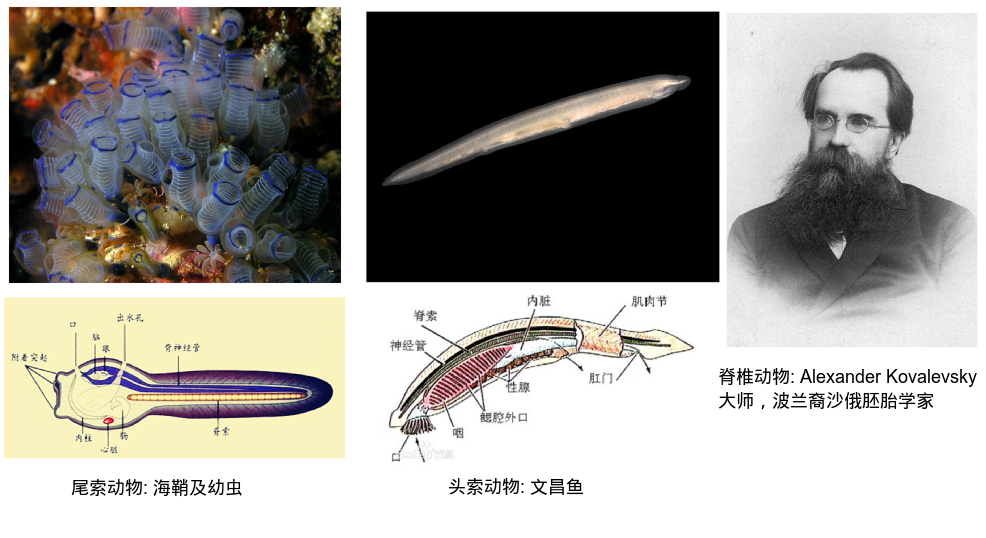
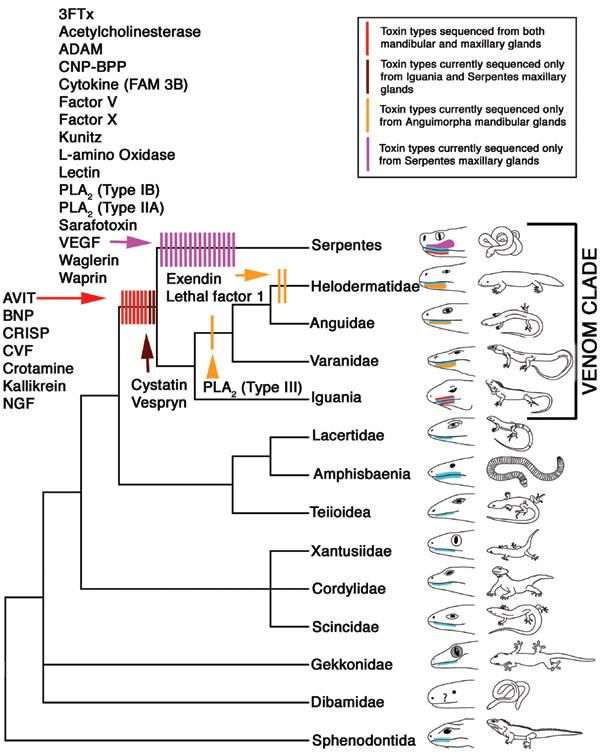
此图credit to 同一篇论文
至于其他的形态学相似之处,则是在相似的环境的自然选择压力之下所产生的趋同演化。
这就涉及到了我在回答开头提到的第二点:DNA测序推动了进化生态学科对自然选择中环境因素的重新认识与思考。
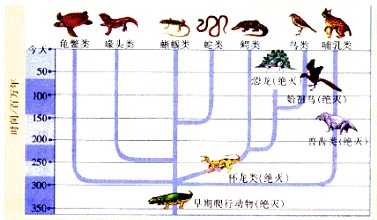
在DNA测序之前,我们并不清楚这样具体的物种之间的系统发生学关系,许多相似的性状被自然理解成物种进化时亲缘关系的承袭。但DNA测序已经用无数例子向我们说明,其实高度相似的性状是可以通过独立演化得到的。诸如 @雪山象回答中提到的水母和栉水母、非洲猬目和猬形目分属不同总目却长得基本一样的刺猬、 @Pepling Yel回答中提到的鬣狗与狗——其实曾经还有一种很象狗的动物叫做塔斯马尼亚虎(Tasmanian tiger),但是它连真哺乳动物亚纲(Eutheria)都不属于,而是一种有袋类动物(和袋鼠、负鼠亲缘关系很近)。
Tasmanian tiger(灭绝于1936年)和澳洲野狗的对比,有没有很像
还有其他许多的例子可举,不过就不展开了。
DNA测序可以真正解读出十分相似的形态背后不同的演化机制,从而让“趋同演化”从一个谈论个例的孤立现象上升成为自然演进中最广泛存在的现象之一。而在认识到趋同演化的普遍性之前,人们难以真正认识到环境因素和对物种基因定向选择的效力到底有强烈。相似的自然选择压力会很容易地使处在相似环境和生态位的物种出现趋同演化;而对比来看,在人们的认识上生物基因的可塑性进一步提高了。
由此,研究生物与周围环境互动关系的生态学和研究生物在地质历史时间里演进的生物进化学的研究视角才正确地得到了整合。继而,我们对于作为整体的生命的可能性与多样性的认识也迈步向前——未尝不能算是自然美学的一种审美意识的进步。
感谢阅读。
注:文中所引用的论文
Fry, B.G. et al. (2006) Early evolution of the venom system in lizards and snakes. Nature 439: 584-588.
图片来自网络
参考文献:
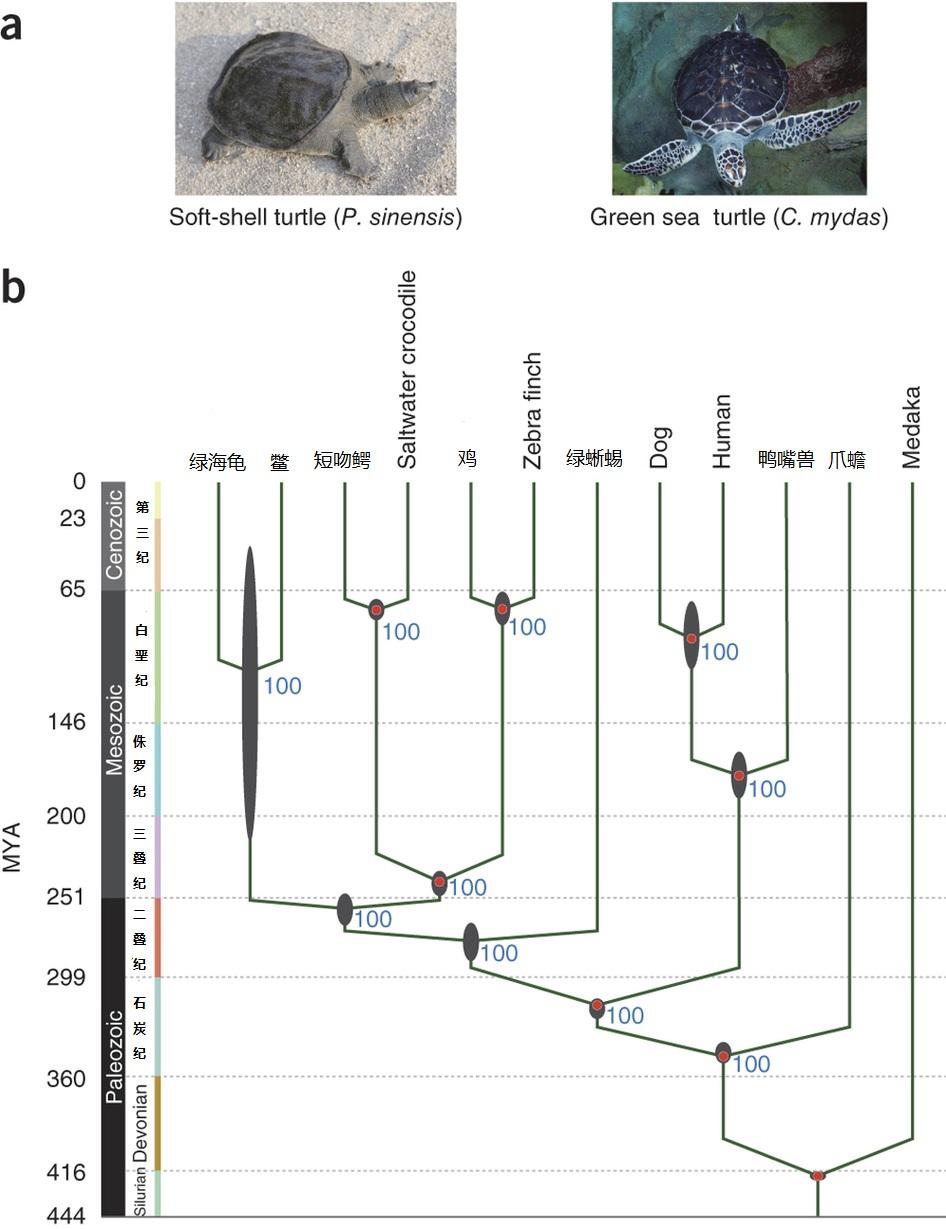
The draft genomes of soft-shell turtle and green sea turtle yield insights into the development and evolution of the turtle-specific body plan. Wang Z, et al. Nat Genet 2013 Jun
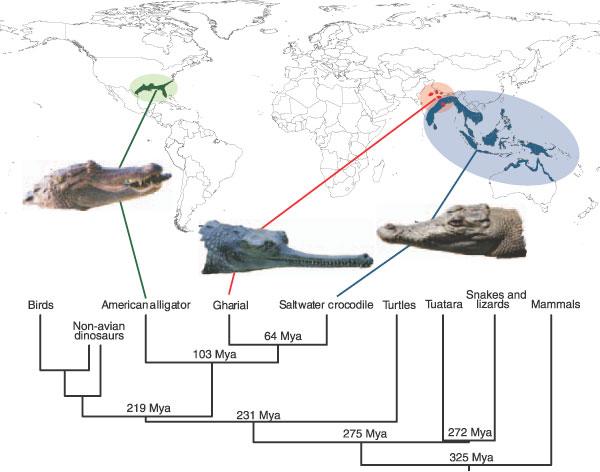
Three crocodilian genomes reveal ancestral patterns of evolution among archosaurs.

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 






 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 发表于 2024-9-1 00:20
发表于 2024-9-1 00:20









 发表于 2024-9-1 00:21
发表于 2024-9-1 00:21
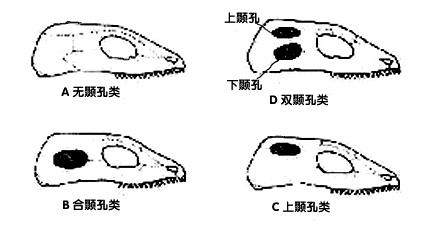
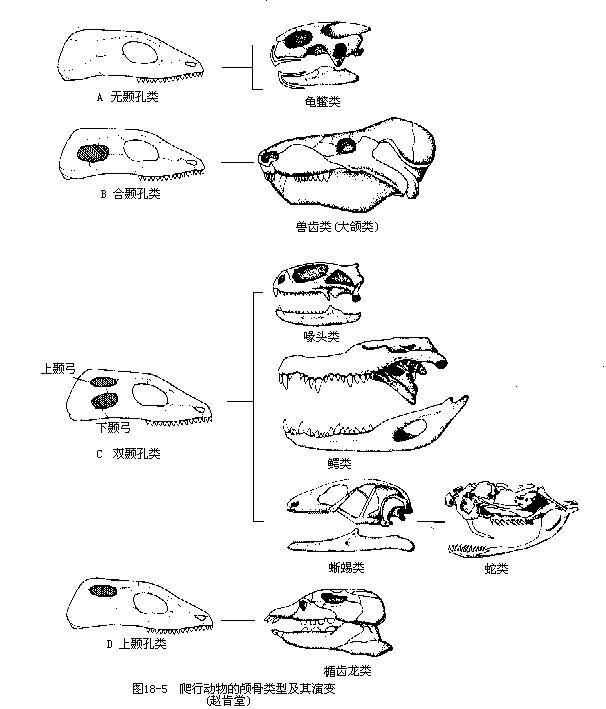




 发表于 2024-9-1 00:22
发表于 2024-9-1 00:22