登陆有奖并可浏览互动!
您需要 登录 才可以下载或查看,没有账号?立即注册


×

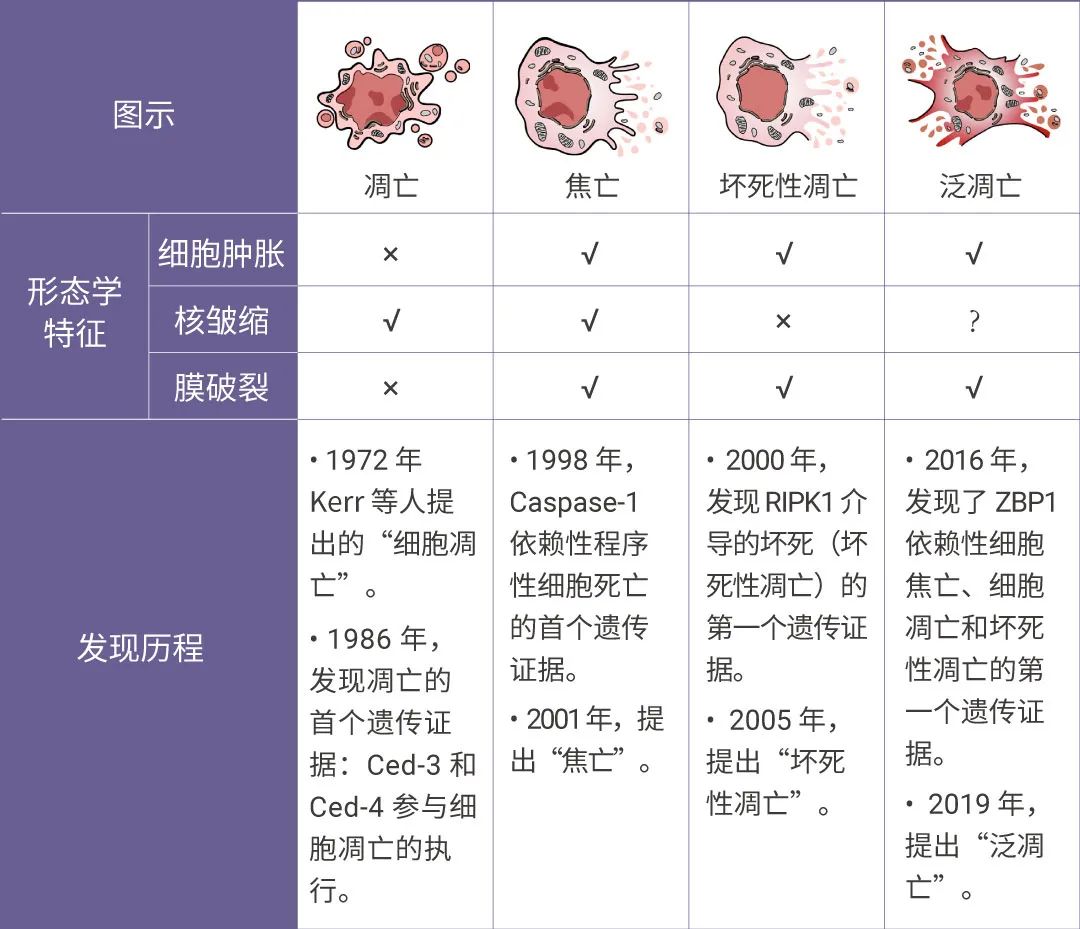
在所有提出的程序性细胞死亡 (PCD) 形式中,细胞焦亡、细胞凋亡和坏死性凋亡是最明确的,其复杂的分子机制负责细胞死亡的启动、转导和执行[1]。但实际上这三种途径之间存在复杂的串扰。例如在无菌损伤和感染性疾病(例如甲型流感病毒 IAV 感染)中,研究人员观察到所有三种 PCD 途径的生化标记物均被激活[1]。
2019 年,美国学者 Malireddi 将这种具有焦亡、凋亡和坏死性凋亡特征的新型死亡方式命名为泛凋亡,并提出先天性免疫传感器 ZBP1 和 TAK1 激酶在泛凋亡小体复合物组装的调控中发挥重要作用[2]。
 Tips: Tips:
泛凋亡(PANoptosis) 是由特定触发器激活并 PANoptosome 复合物调节的炎症性 PCD 途径,其综合了细胞焦亡、细胞凋亡和/或坏死性凋亡的关键特征,这也是 PANoptosis 术语中 “P” “A” 和 “N” 的来源,但其不能被细胞焦亡、凋亡和坏死性凋亡中任意一种死亡方式单独表征[1]。
图 1. 程序性细胞死亡的研究时间表和特征[1]。
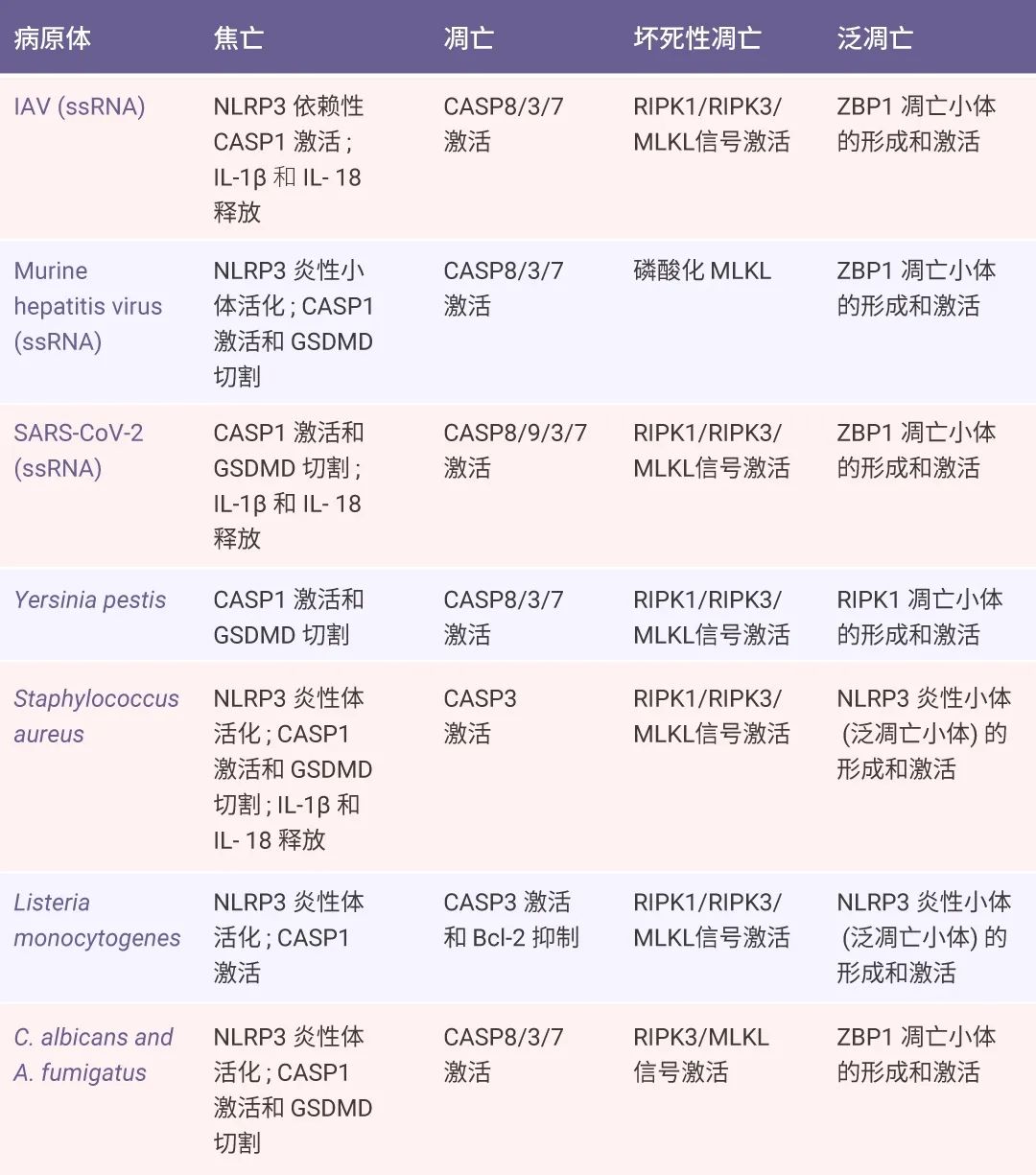
细胞死亡对于机体正常发育和抵抗病原体入侵至关重要,是宿主应对病原感染的一种防御手段。研究发现,IAV 感染后和 TAK1 活性丧失后会发生泛凋亡。此外,多种病原体如病毒、细菌、真菌,甚至寄生虫,以及其他非感染性因素如肿瘤中的细胞因子,都可能引发宿主细胞的泛凋亡 (表 1)[3]。表 1. 可诱导巨噬细胞焦亡、凋亡和坏死的病原体[3]。

多种蛋白质可以形成调节 PCD 的多蛋白复合物,其基于各种蛋白质结构域之间的相互作用,可分为三类:传感结构域、组装结构域和催化结构域[2]。
▐泛凋亡小体: PANoptosome
泛凋亡受到上游感受器和分子信号的级联调控,这些传感器和级联信号组装成一种多聚体复合物,即泛凋亡小体 (PANoptosome)。PANoptosome 及其上游的感受器不仅是下游分子的激活平台,也是 3 种 PCD 途径的启动“总开关”[4][5]。
PANoptosome 小体作为一种分子支架,允许参与焦亡、细胞凋亡和/或坏死性凋亡的关键分子进行耦联和结合[1]。传感器蛋白在感知到病原体成分后,会介导 RIPK3、RIPK1、CASP8、FADD等蛋白组装成 PANoptosome 复合体,从而诱导 PANoptosis[2][6]。
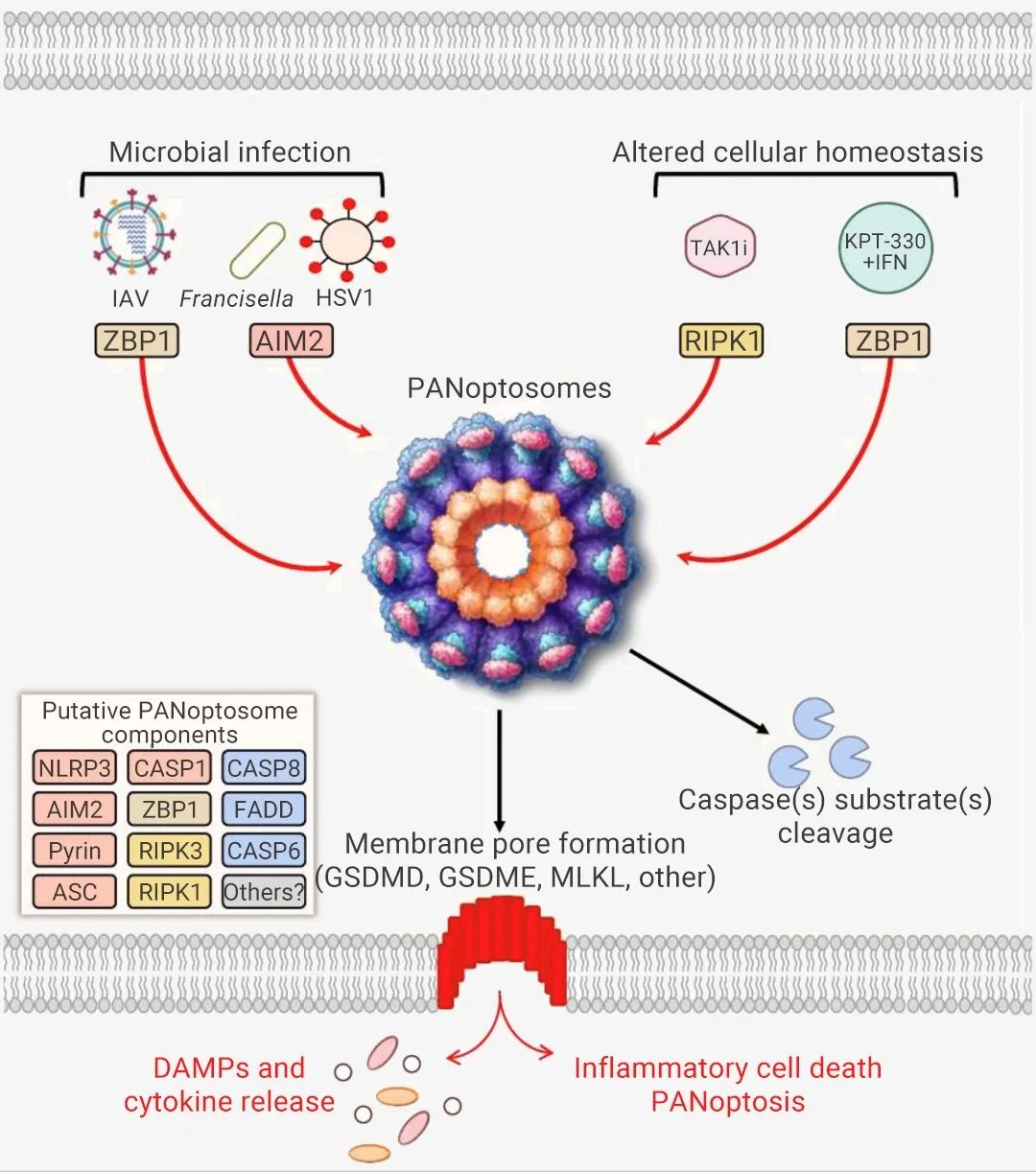
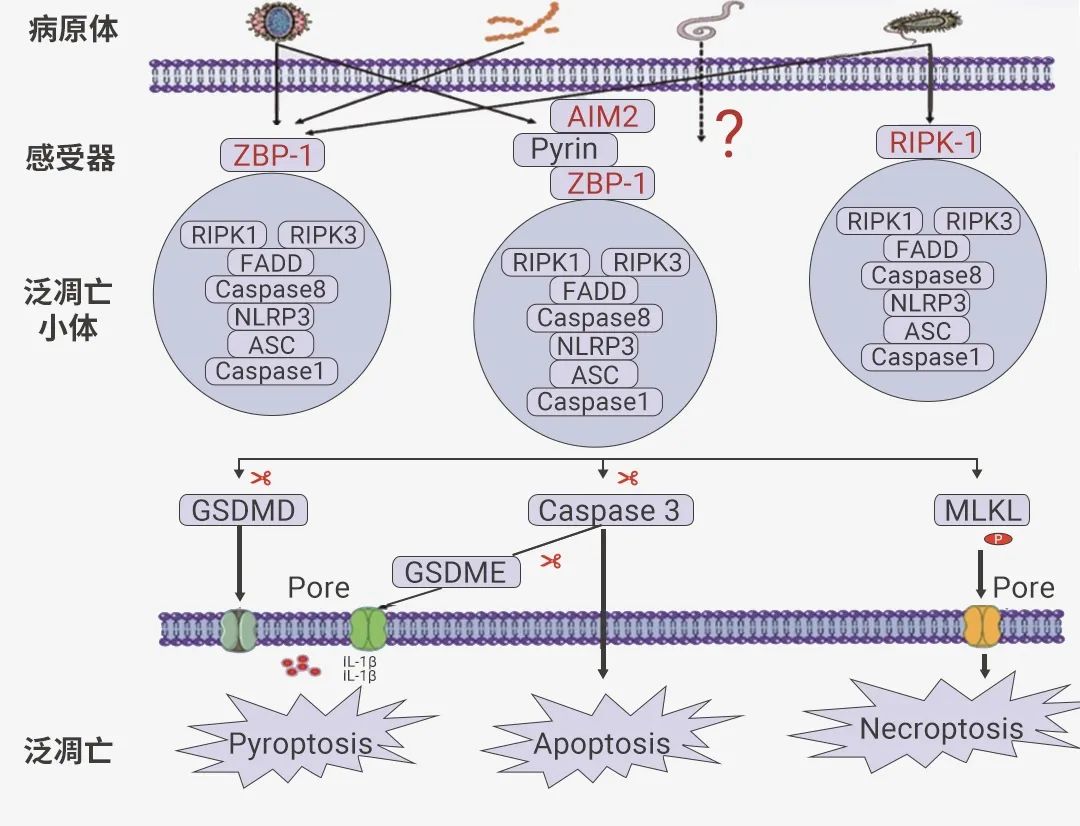
构成 PANoptosome 小体的蛋白质通常可分为三类:(1) ZBP1 和 NLRP3 作为假定的 PAMP 和 DAMP 传感器,(2) ASC 和 FADD 作为适配器,以及 (3) RIPK1、RIPK3 、CASP1 和 CASP8 作为催化效应器[2]。图 2. PANoptosis 和 PANoptosome 形成[1]。
那么上游感受器如何特异性识别病原微生物感染以及这些成分之间相互作用的呢?其实这个的具体机制目前还是未知的。迄今为止,已经明确的泛凋亡上游分子有三种,分别是 ZBP1、RIPK1 和 AIM2,它们可以感受特定刺激并触发泛凋亡小体的组装,形成三种具有不同传感器和调节因子的泛凋亡小体,即 ZBP1-PANoptosomes、AIM2-PANoptosomes 和 RIPK1-PANoptosomes。此外,PANoptosome 的激活也会受到 TAK1、PSTPIP2、SHARPIN、HOIP、HOIL-1 和 A20 的抑制[6]。图 3. 泛凋亡小体及其通路[4]。
 Tips: Tips:
(1) ZBP1 泛凋亡小体:Z-DNA 结合蛋白 1 (ZBP1) 是一种先天免疫受体,被鉴定为甲型流感病毒(Influenza A virus,IAV)的感受器和 IAV 感染期间细胞死亡的主要调节因子。在 IAV 刺激下,ZBP1 作为特异的感受器被激活,进而触发 PANoptosome 的组装[7][8]。ZBP1-PANoptosis 形成后,可以启动由焦亡、凋亡和坏死性凋亡成分组成的泛凋亡,最终导至溶解性炎症细胞死亡,其特征是 Caspase-1、Caspase-3、Caspase-8 的激活和 MLKL 的磷酸化[9]。
(2) RIPK1 泛凋亡小体:RIPK1 介导的泛凋亡小体主要由 RIPK1、ASC、Caspase-1、Caspase-8 和死亡相关结构域蛋白(fas-associated protein with death domain,FADD)构成,最早由 Malireddi 教授团队提出。该团队还发现 TAK1 可作为 RIPK1-PANoptosome 的调控主开关,因为 TAK1 基因的缺失或者功能失活可以触发 RIPK1-PANoptosome 的组装。然而,后续研究发现,TAK1 的缺乏同样可以诱导 RIPK1 非依赖的泛凋亡通路的激活,后者主要由 RIPK3-MLKL 介导。
(3) AIM2 泛凋亡小体:其组成分子包括 AIM2、ZBP1、pyrin、ASC、Caspase-1、Caspase-8、RIPK1、RIPK3 和 FADD[10]。AIM2 炎性小体可感知双链 DNA,AIM2 调节先天免疫传感器 Pyrin 和 ZBP1 来驱动炎症信号传导和炎症细胞泛凋亡,在人类的正常发育、传染性疾病、炎症和肿瘤方面发挥重要作用。

说了这么多,这种 “三合一” 的复杂细胞死亡方式,又该如何研究及检测的呢?
首先咱们来说检测,由于泛凋亡综合了细胞焦亡、细胞凋亡和/或坏死性凋亡的关键特征,因此在进行相关检测时三方面缺一不可!
之前小 M 也为大家整理过相关的检测方法,那么本期仅做整理,不再详述。
 Tips:泛凋亡检测方法及指标! Tips:泛凋亡检测方法及指标!
(1) 观察细胞形态:细胞焦亡引起细胞质肿胀和膜破裂;凋亡主要形态学特征包括染色质固缩、DNA 片段化、细胞膜起泡、细胞皱缩、凋亡小体的形成等。
(2) 检测不同 PCD 的关键蛋白:焦亡相关:Caspase-1、Caspase-3、Gasdermins、AIM2/Pyrin/NLRP3 等;凋亡相关:Caspase-3、Caspase-7、Caspase-8、PARP、Bax/Bcl 等;坏死性凋亡相关:MLKL、RIPK1、RIPK3、ZBP1 等。
(3) 其他指标检测:Annexin V-FITC 和 PI 联合染色;TUNEL 法;JC-1 检测;ELISA 检测炎症因子的释放;免疫印迹、流式等技术检测炎性小体 NLRP3 的表达和 Caspase-1 的活化情况。
▐文献案例解读(IF=39.3):
近期,Jin-Fei Lin 等人发现磷酸化的 NFS1 可通过预防 PANoptosis 以削弱结直肠癌的 Oxaliplatin 敏感性[11]。
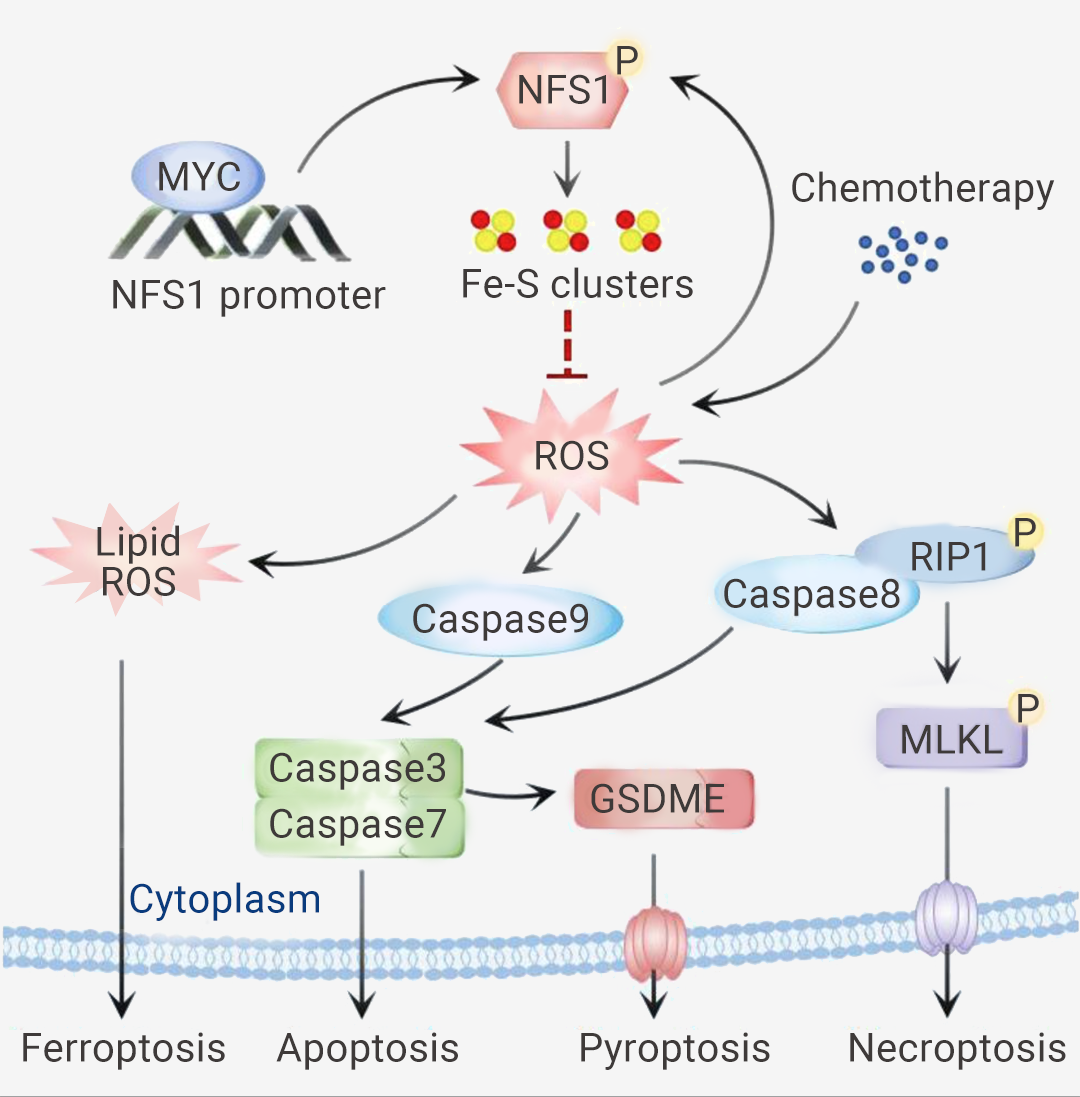
作者使用基于体内代谢酶基因的 (CRISPR)-Cas9 文库筛选,发现 NFS1 的缺失显著增强了 CRC 细胞对奥沙利铂的敏感性。体外和体内结果表明,NFS1 缺陷与 Oxaliplatin 协同作用,通过增加细胞内活性氧 (ROS) 水平,引发泛凋亡。图 4. 磷酸化的 NFS1 通过降低 ROS 水平来减弱铂基化学敏感性,从而防止 PANoptosis[11]。
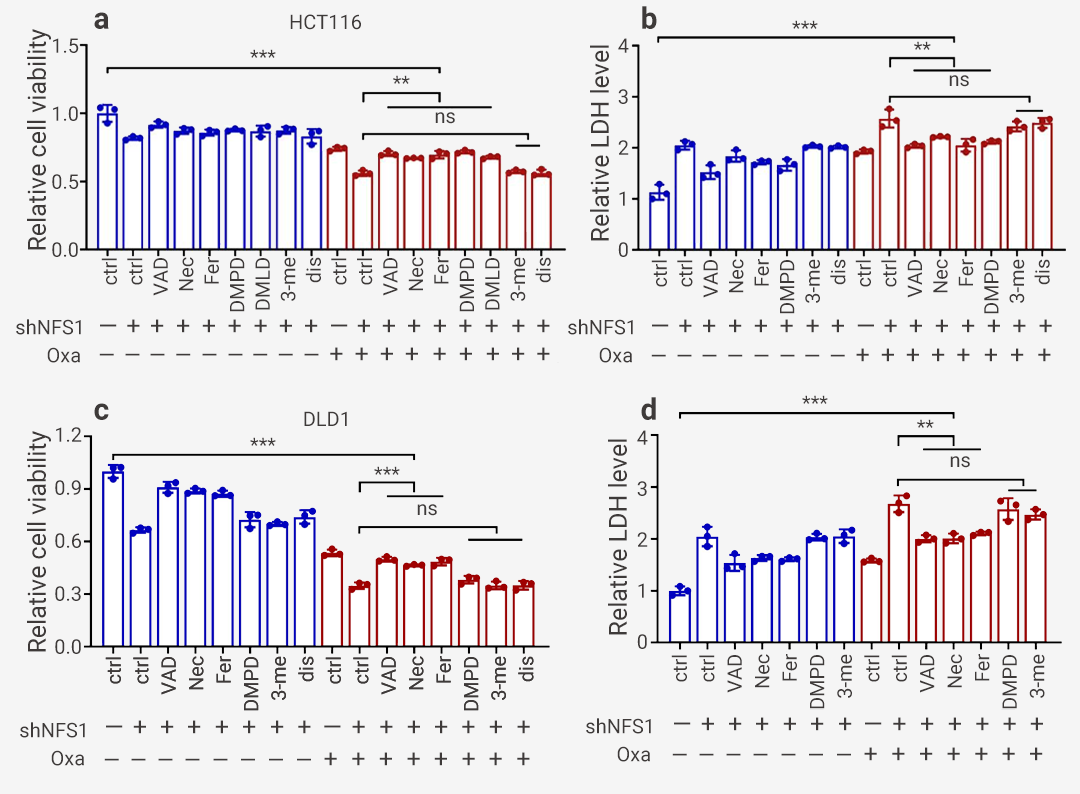
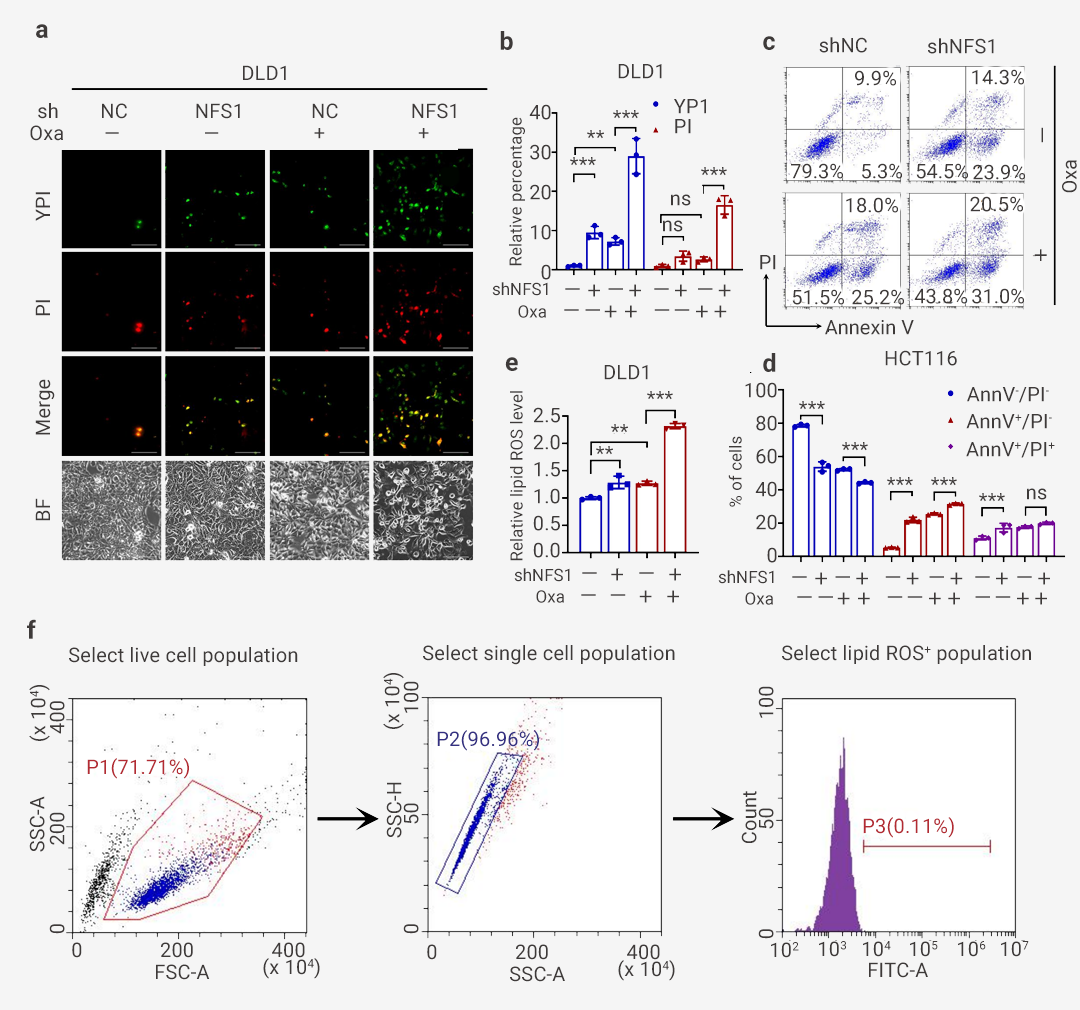
为了研究发生的细胞死亡类型,使用常见细胞死亡途径的多种抑制剂并结合光镜观察细胞形态、YP1/PI 染色、流式检测细胞死亡等进行检测。结果表明,焦亡 (GSDME) 抑制剂 Disulfiram 和自噬抑制剂 3-methyladenine 无显著作用,而凋亡抑制剂 Z-VAD-FMK 、坏死性凋亡抑制剂 Necrostatin-1 、铁死亡抑制剂 Ferrostatin-1 、焦亡 (GSDME) 抑制剂 Ac-DMPD/DMLD-CMK 逆转(但不能完全恢复)Oxaliplatin 治疗下由 NFS1 缺乏引起的细胞活力降低和细胞毒性增强 (图 5)。图 5. Oxaliplatin 联合多种抑制剂的细胞活力及毒性测定[11]。 a-b: 对照和 NFS1 敲低 HCT116 细胞的细胞活力 (a) 和细胞毒性 (b) 评估; c-d: 对照和 NFS1 敲低 DLD1 细胞活力 (c) 和细胞毒性 (d) 评估。
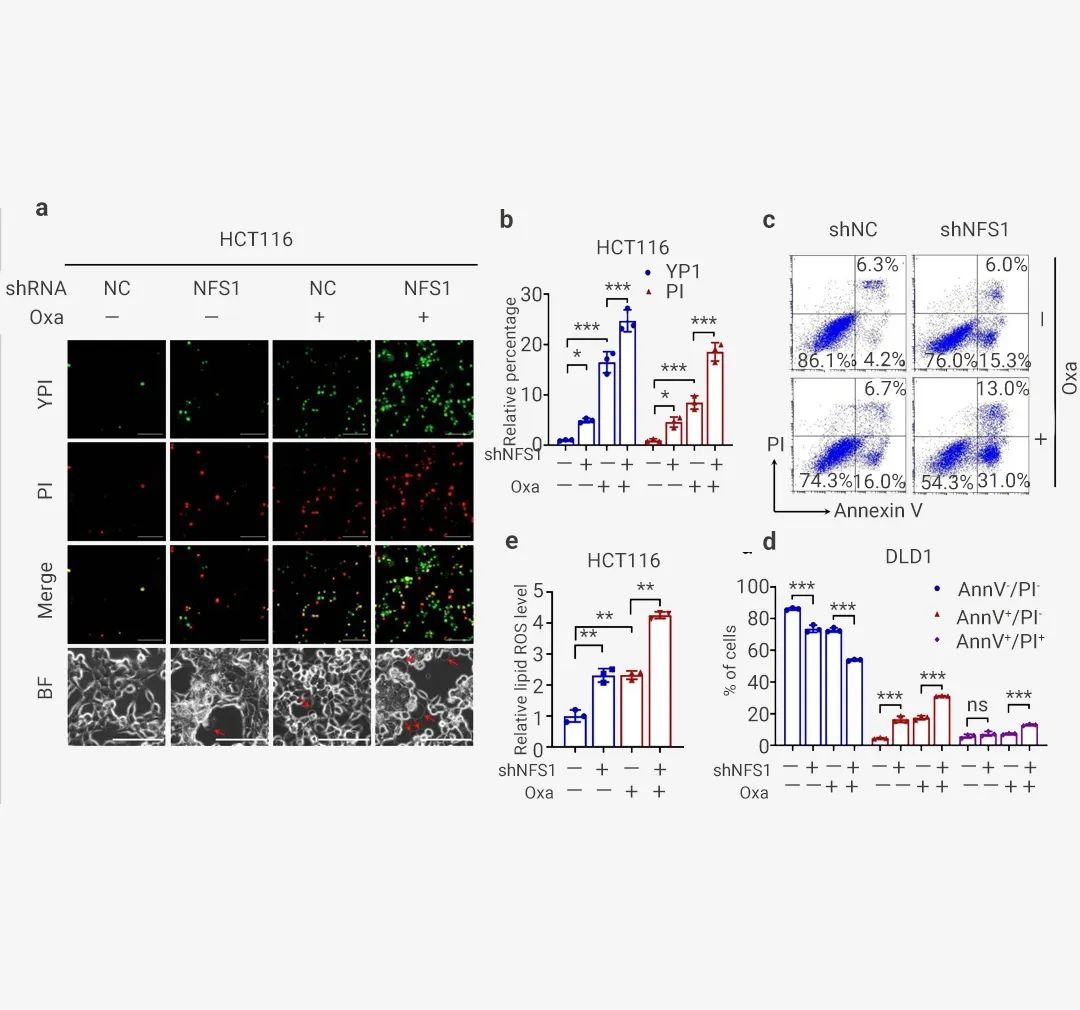
为了进一步证实 PANoptosis 的发生,作者发现 NFS1 敲低联合 Oxaliplatin 治疗显著增加了死亡细胞的数量,包括指示细胞凋亡或坏死性凋亡的 YP1 阳性细胞和指示细胞坏死性凋亡、细胞焦亡或铁死亡的 PI 阳性细胞 (图 6: 左图, a-b 和右图 a-b)。
对 HCT116 细胞的分析还表明,联合组中从质膜出现的大气泡数量明显高于阴性对照组、NFS1 敲低组和 Oxaliplatin 治疗组,这表明细胞焦亡的发生 (图 6: 左图, a)。此外,NFS1 耗竭后进行 Oxaliplatin 治疗定量增加了早期/晚期凋亡细胞的数量,并显著减少了活细胞的数量 (图 6: 左图, c-d 和右图 c-d)。此外,在 NFS1 敲低组和 Oxaliplatin 治疗组中检测到的脂质 ROS 含量高于对照组。这种增加在联合组中更为明显,表明铁死亡的发生 (图 6: 左图, e 和右图 e-f)。因此,这些数据表明 NFS1 缺陷与 Oxaliplatin 联合有助于 PANoptosis 的激活。
(左图): a. 对照中可能发生凋亡或坏死性凋亡的 YP1+细胞(绿色)和可能发生凋亡、坏死性凋亡、细胞焦亡或铁死亡的 PI+细胞(红色)以及用 Oxaliplatin 处理的 NFS1 敲低 HCT116 细胞。底部显示为明场,红色箭头表示从质膜中出现的大气泡; b. 对 (a) 中的 YP1+ 和 PI+ 细胞进行定量。c-d: 流式细胞术 (c) 和采用膜联蛋白 V/PI 染色的定量分析 (d),在对照和用 PBS 或 Oxaliplatin 处理的 NFS1 敲低 DLD1 细胞中评估活细胞 (Annexin V- /PI-)、早期凋亡细胞 (Annexin V+ /PI-) 和晚期凋亡细胞的百分比 (Annexin V+ /PI+); e. 通过 BODIPY™ 581/591 C11 探针测定评估用 Oxaliplatin 处理的对照和 NFS1 敲低 HCT116 细胞中的脂质 ROS 水平。
(右图): a. 对照中可能发生凋亡或坏死性凋亡的 YP1+细胞(绿色)和可能发生凋亡、坏死性凋亡、细胞焦亡或铁死亡的 PI+ 细胞(红色)以及用 Oxaliplatin 处理的 NFS1 敲低 DLD1 细胞。底部显示为明场; b. 对 (a) 中的 YP1+ 和 PI+ 细胞进行定量; c. 使用膜联蛋白 V/PI 染色进行流式细胞术分析,评估对照和用 PBS 或 Oxaliplatin 处理的和 NFS1 敲除 HCT116 中活细胞 (Annexin V-/PI-)、早期凋亡细胞 (Annexin V+/PI-) 和晚期凋亡细胞 (Annexin V+/PI+) 的百分比; d. (c)中活细胞 (Annexin V-/PI-)、早期凋亡细胞 (Annexin V+/PI-) 和晚期凋亡细胞 (Annexin V+/PI+) 的定量; e. 使用 BODIPYTM 581/591 C11 探针测定法评估Oxaliplati 处理的对照细胞和 NFS1 敲低 DLD1 细胞中的脂质 ROS; F. 本研究中用于 ROS/脂质 ROS 分析的策略。
此外,作者评估了 Oxaliplatin 治疗下 NFS1 缺陷诱导 PANoptosis 的具体机制。总之,Oxaliplatin 介导的氧化应激可增强 NFS1 的丝氨酸磷酸化水平,并且 NFS1 以 S293 磷酸化依赖性方式阻止 Oxaliplatin 治疗下 PANoptosis 的激活。
本期小 M 为大家介绍了细胞死亡的新姿势:泛凋亡 (PANopotosis),一种炎症性细胞程序死亡途径。三种重要的泛凋亡小体 ZBP1-PANoptosomes、AIM2-PANoptosomes 和 RIPK1-PANoptosomes 在其中发挥重要的调节作用,此外,小 M 为大家介绍了泛凋亡的检测指标,并通过文献案例进一步详述。希望大家能够有所收获喔~
MCE的所有产品仅用作科学研究或药证申报,我们不为任何个人用途提供产品和服务。
参考文献:
[1] Wang Y, et al. From pyroptosis, apoptosis and necroptosis to PANoptosis: A mechanistic compendium of programmed cell death pathways. Comput Struct Biotechnol J. 2021 Aug 3;19:4641-4657.
[2] Malireddi RKS, et al. ZBP1 and TAK1: Master Regulators of NLRP3 Inflammasome/Pyroptosis, Apoptosis, and Necroptosis (PAN-optosis). Front Cell Infect Microbiol. 2019 Nov 26;9:406.
[3] Zhu P, et al. Advances in mechanism and regulation of PANoptosis: Prospects in disease treatment. Front Immunol. 2023 Feb 9;14:1120034.
[4] Pengyue Zhao, et al. Research progress on the role of PANoptosis in human diseases. August 2022Zhonghua Yi Xue Za Zhi 102(32):2549-2554.
[5] Christgen S, et al. Identification of the PANoptosome: A Molecular Platform Triggering Pyroptosis, Apoptosis, and Necroptosis (PANoptosis). Front Cell Infect Microbiol. 2020 May 29;10:237.
[6] Samir P, et al. The PANoptosome: A Deadly Protein Complex Driving Pyroptosis, Apoptosis, and Necroptosis (PANoptosis). Front Cell Infect Microbiol. 2020 Jun 3;10:238.
[7] Gullett JM, et al. It's All in the PAN: Crosstalk, Plasticity, Redundancies, Switches, and Interconnectedness Encompassed by PANoptosis Underlying the Totality of Cell Death-Associated Biological Effects. Cells. 2022 Apr 29;11(9):1495.
[8] Zheng M, et al. Newly Identified Function of Caspase-6 in ZBP1-mediated Innate Immune Responses, NLRP3 Inflammasome Activation, PANoptosis, and Host Defense. J Cell Immunol. 2020;2(6):341-347.
[9] Zheng M, et al. The regulation of the ZBP1-NLRP3 inflammasome and its implications in pyroptosis, apoptosis, and necroptosis (PANoptosis). Immunol Rev. 2020 Sep;297(1):26-38.
[10] Lee S, et al. AIM2 forms a complex with pyrin and ZBP1 to drive PANoptosis and host defence. Nature. 2021 Sep;597(7876):415-419.
[11] Lin JF, et al. Phosphorylated NFS1 weakens oxaliplatin-based chemosensitivity of colorectal cancer by preventing PANoptosis. Signal Transduct Target Ther. 2022 Feb 28;7(1):54.
|

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 





 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 雷达卡
雷达卡 发表于 2024-1-30 13:58
发表于 2024-1-30 13:58



 提升卡
提升卡


