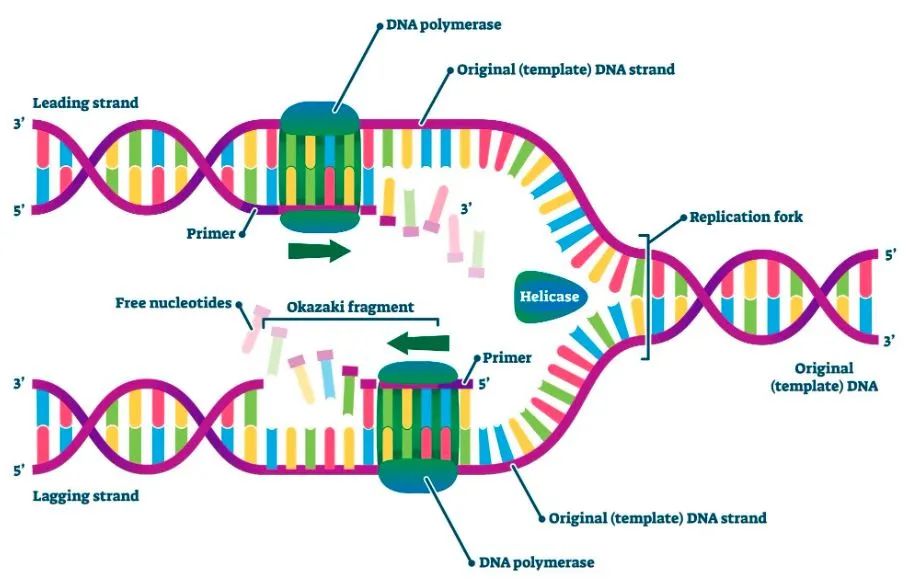
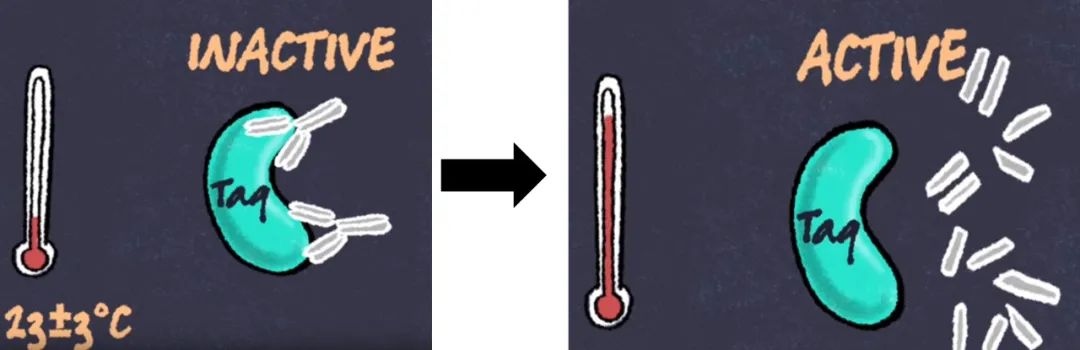
DNA 聚合酶因其在合成新 DNA 链中的关键作用而成为 PCR 的重要组成部分。因此,了解这种酶的特性以及先进 DNA 聚合酶的后续开发对于将 PCR 的功能应用于广泛的生物应用至关重要。 自从在早期 PCR 检测中使用Taq DNA 聚合酶以来,PCR 酶的特异性、热稳定性、保真性和持续合成能力得到了显著改进,以增强PCR扩增的性能,下文详述Taq聚合酶的这几个特性: 非特异性扩增是 PCR 的主要障碍之一,非常影响目标扩增的产量和灵敏度,从而影响PCR扩增结果和下游应用的成功。DNA 聚合酶通常扩增错误引物的目标和引物二聚体,这些是常见的非特异性扩增。 减少非特异性扩增的一种方法是在冰浴上配置PCR反应,有助于保持 DNA 聚合酶的活性在较低的水平,但在 PCR 开始之前仍可能会合成不需要的产物。 另一种解决方案是延迟添加 DNA 聚合酶,直到第一个循环的退火步骤才添加。这种技术被称为“热启动”,因为只有在 90°C 以上的初始变性步骤后才能开始扩增。 尽管可有效提高特异性,但手动热启动过程费力且会增加样品污染和重现性差的风险。1994 年,开发出具有真正热启动特性的Taq DNA 聚合酶 [1,2],将特异性抗体与聚合酶结合,在配置反应体系时可抑制聚合酶的活性。在初始高温变性步骤(例如,>90°C)中,结合的抗体被降解,从而激活 DNA 聚合酶(图 1)。
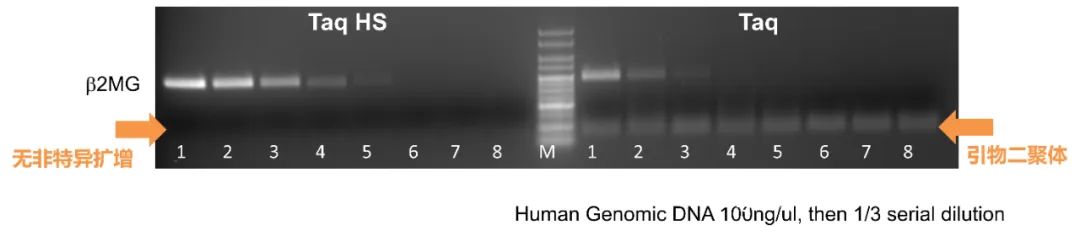
图 1. 基于抗体的热启动 DNA 聚合酶 变性步骤还分离了在反应设置过程中可能形成的错误引物和引物二聚体,从而防止它们在随后的退火和延伸步骤中被 DNA 聚合酶扩增。以这种方式,热启动 DNA 聚合酶减少了非特异性扩增,增加了产量,并为高通量应用提供了方便的室温设置。
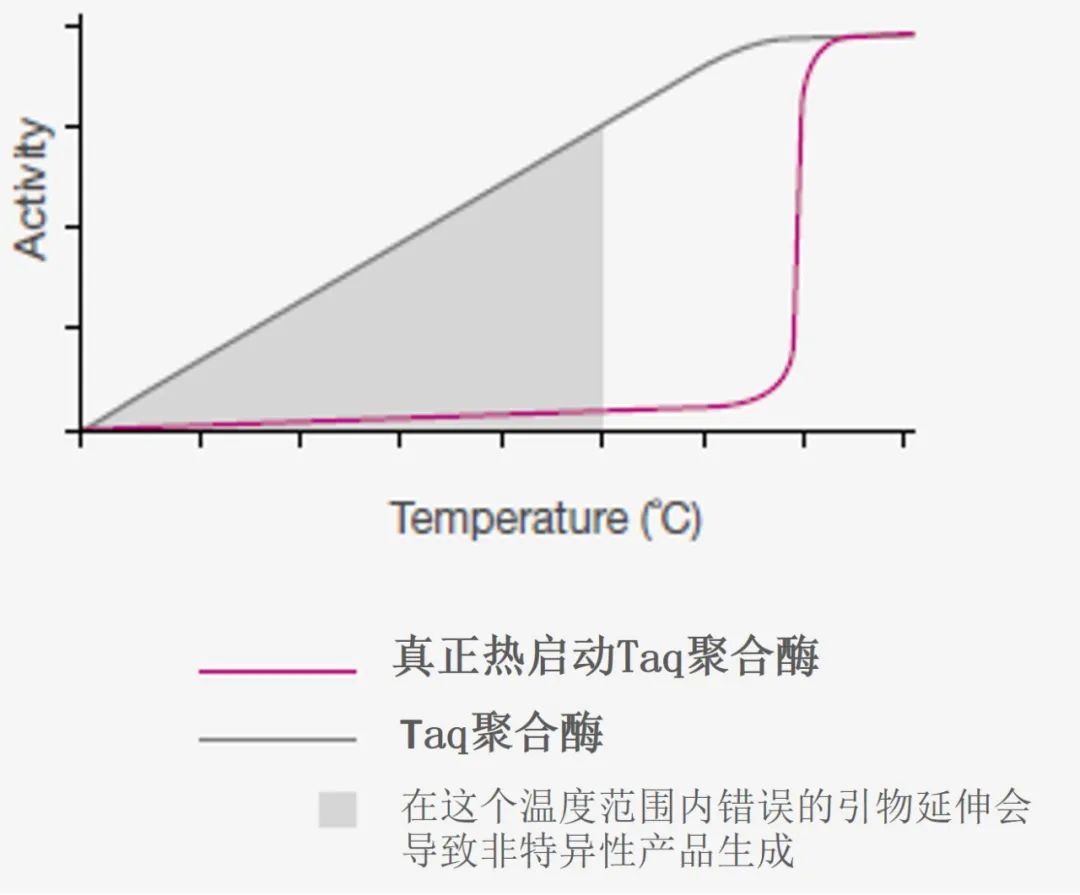
图 2. 聚合酶活性比较: 热启动DNA Taq 聚合酶与普通Taq 聚合酶。真正的热启动Taq聚合酶在 95°C 下热激活后,抗体从聚合酶中解离才可显示出活性,而普通的Taq 聚合酶在加热过程中依然有活性,因此会导至非特异性产物生成。
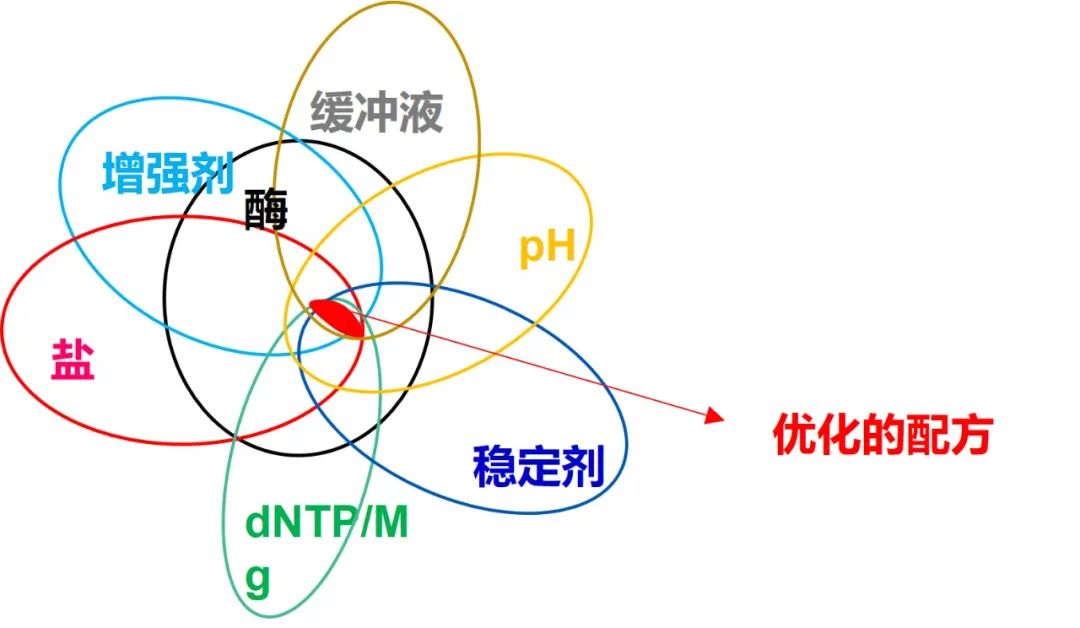
作为抗体的替代品,热启动属性也可以通过化学修饰以及适配体修饰(缩短激活时间)来实现。无论选择何种热启动技术,DNA 聚合酶的活性在未加热条件下被有效阻断以确保特异性至关重要(图 2)。 由于热循环是实现扩增 DNA 重复链式反应的条件的关键特征,因此所使用的 DNA 聚合酶的热稳定性是一个重要特征。 虽然最初源自嗜热细菌菌株的Taq DNA 聚合酶可以承受相对较高的温度,但其半衰期在 90°C 以上时显著缩短。当使用长时间的高温使具有二级结构和富含 GC 序列的 DNA 变性时,这种不足会带来挑战。 同样,在扩增长模板时,TaqDNA 聚合酶可能需要以更高的量提供或补充以延长孵育时间。因此,从超嗜热生物体中分离出的 DNA 聚合酶由于其较高的热稳定性,已成为克服这些挑战的工具。 一种众所周知的超热稳定酶是Pfu DNA 聚合酶,来自热液环境中发现的古菌超嗜热火球菌。在 95°C 下,Pfu聚合酶的稳定性比Taq聚合酶稳定约 20 倍 [3]。其他流行的超热稳定 DNA 聚合酶包括来自古菌Thermococcus和Pyrococcus物种的 KOD 和 GBD 。 尽管古细菌 DNA 聚合酶具有极高的热稳定性,但它们在某些情况下可能存在局限性。例如,由于较低的持续合成能力(与Taq DNA 聚合酶相比),超热稳定Pfu DNA 聚合酶合成 DNA 的速度较慢。 此外,由于尿嘧啶结合口袋作为 DNA 修复机制的存在,古细菌 DNA 聚合酶无法扩增含有尿嘧啶的 DNA 模板 [4,5]。含尿嘧啶的 DNA 序列是通过亚硫酸氢盐转化进行PCR 残留预防和基因座甲基化分析的基础。 DNA 聚合酶的校对能力决定了保真度,从而提高了 DNA 序列复制的准确性。高保真DNA聚合酶是具有很强校对活性的酶。DNA 聚合酶准确复制 DNA 序列(即获得低错误序列)的能力在克隆、测序和定点诱变等应用中至关重要。 DNA 聚合酶的校对活性基于其 3' → 5' 核酸外切酶活性,可纠正错误掺入的核苷酸。核酸外切酶活性发生在 DNA 聚合酶上与其 5'→ 3' 聚合酶活性位点分开的位置。 当不匹配的核苷酸掺入聚合域时,DNA 合成会因不利的碱基配对动力学而停滞。延迟允许切除不匹配的核苷酸,并通过 DNA 聚合酶用正确的核苷酸替换它 [6]。 DNA 聚合酶的保真度通常表示为错误率的倒数(保真度 = 1/错误率),这是指错误掺入的核苷酸数与聚合的核苷酸总数之比。因此,测得的 DNA 聚合酶保真度在很大程度上取决于 PCR 扩增子的长度以及用于生成 PCR 产物的 PCR 循环数。 通常,保真度表示为相对于Taq DNA 聚合酶的保真度。天然存在的校对 DNA 聚合酶(例如Pfu和 KOD)的保真度约为Taq保真度的 10 倍。然而,“下一代”高保真 DNA 聚合酶是通过定向进化设计的,具有出色的保真度,是Taq聚合酶的 50-300 倍以上。对于这些酶,错误掺入率约为百万分之一的掺入核苷酸。 酶的持续合成能力定义为在单个结合事件中被处理的核苷酸数量。DNA 聚合酶的持续合成能力通常反映合成速率和速度,以及对其底物的亲和力。因此,高加工能力的 DNA 聚合酶有利于扩增长模板和具有二级结构和高 GC 含量的序列,并且存在血液和植物组织中存在的 PCR 抑制剂(例如肝素、木聚糖和腐殖酸)。 早期的高保真 DNA 聚合酶往往表现出较低的持续合成能力,因为它们具有很强的核酸外切酶活性,从而减慢了聚合反应。因此,长靶标 DNA 的扩增速度会显著减慢。例如,校对Pfu DNA聚合酶的保真度是Taq DNA聚合酶的7倍,但其合成率不到Taq聚合酶的一半。 当 DNA 聚合酶被设计成具有另一种蛋白质的强 DNA 结合结构域而不影响聚合酶活性时,持续合成能力取得了突破[10]。这种工程化 DNA 聚合酶的持续合成能力增强了 2 至 5 倍。 DNA 聚合酶的四个特性(特异性、热稳定性、保真度和持续合成能力)共同使这些酶具有高度通用性,随后的增强功能进一步拓宽了它们在 PCR 中的应用。 酶的特异性确保以高产量获得所需的 PCR 产物,同时最大限度地减少克隆和定量等下游应用中的潜在问题。 强大的持续合成能力和超热稳定性克服了二级结构、富含 GC 的序列和长 DNA 扩增的困难。此外,改进的持续合成能力使 DNA 样品对天然存在的 PCR 抑制剂具有抗性。 最后是高保真提供序列复制的准确性。 更值得一提的是,决定PCR扩增性能的并不只是酶,整个体系的缓冲液配方(盐,pH, Mg,稳定剂,加强剂,核苷酸等)的每一个组分都会影响反应的性能。只有当各组分优化地搭配在一起,才能发挥出酶的最大活性。
参考文献:
|

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号